

The Frequency of Vaccine-Induced T-Cell Responses Does Not Predict the Rate of Acquisition after Repeated Intrarectal SIVmac239 Challenges in Mamu-B*08+ Rhesus Macaques
- PMID: 30541854
- PMCID: PMC6384082
- DOI: 10.1128/JVI.01626-18
The Frequency of Vaccine-Induced T-Cell Responses Does Not Predict the Rate of Acquisition after Repeated Intrarectal SIVmac239 Challenges in Mamu-B*08+ Rhesus Macaques
Abstract
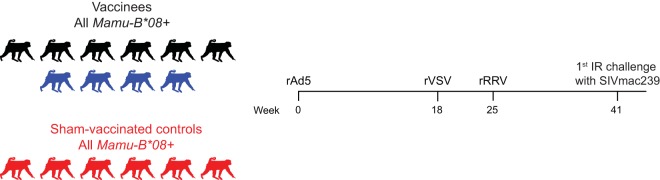
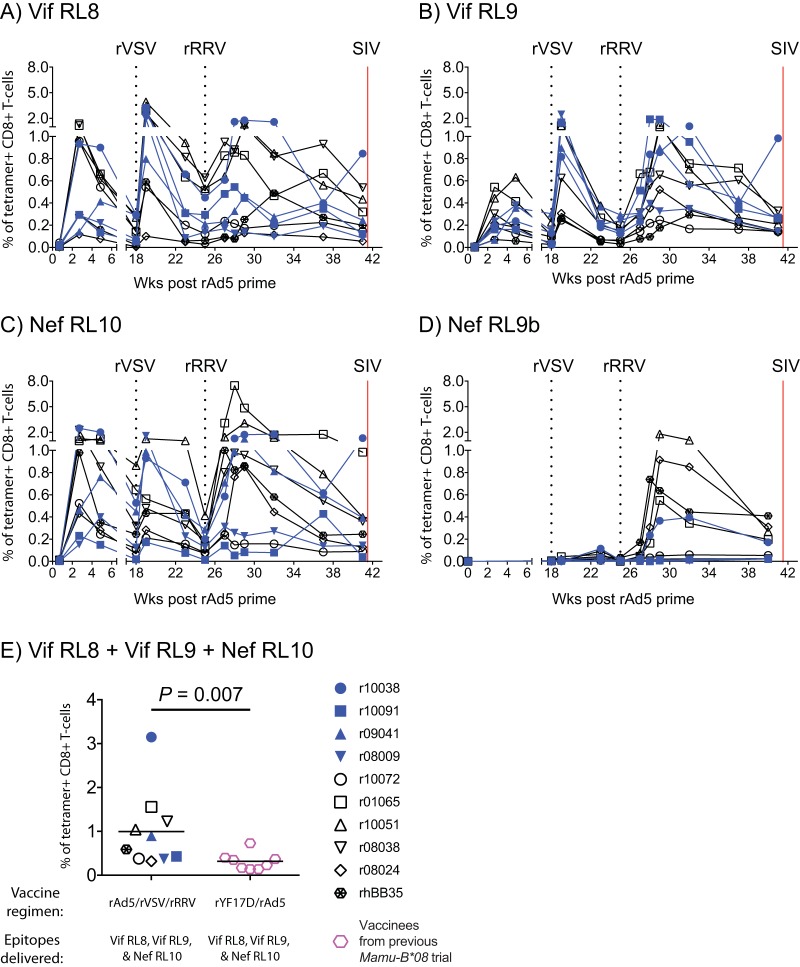
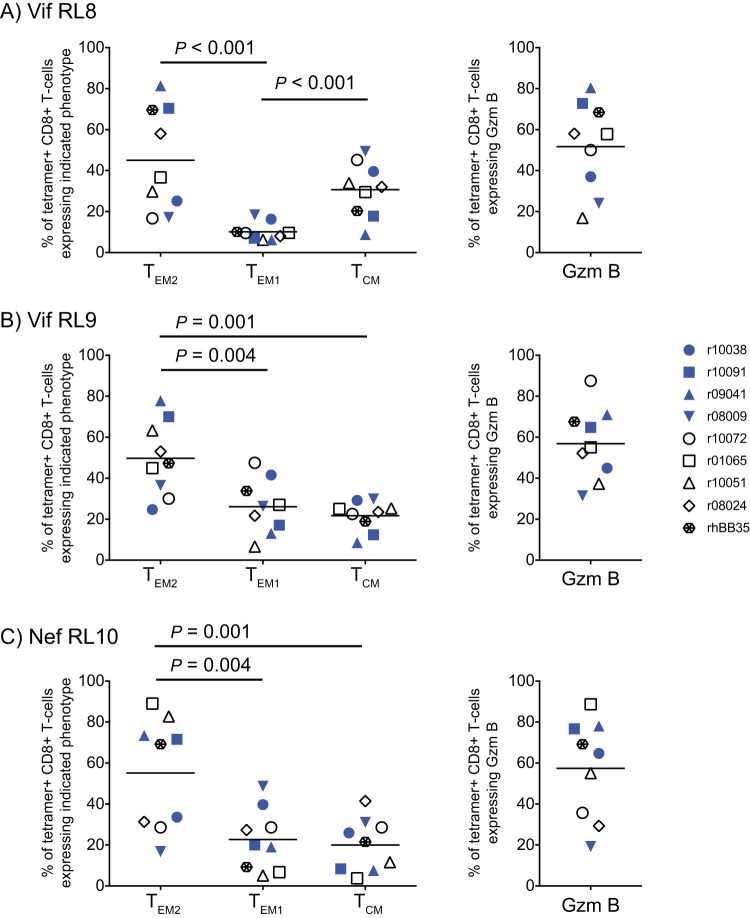
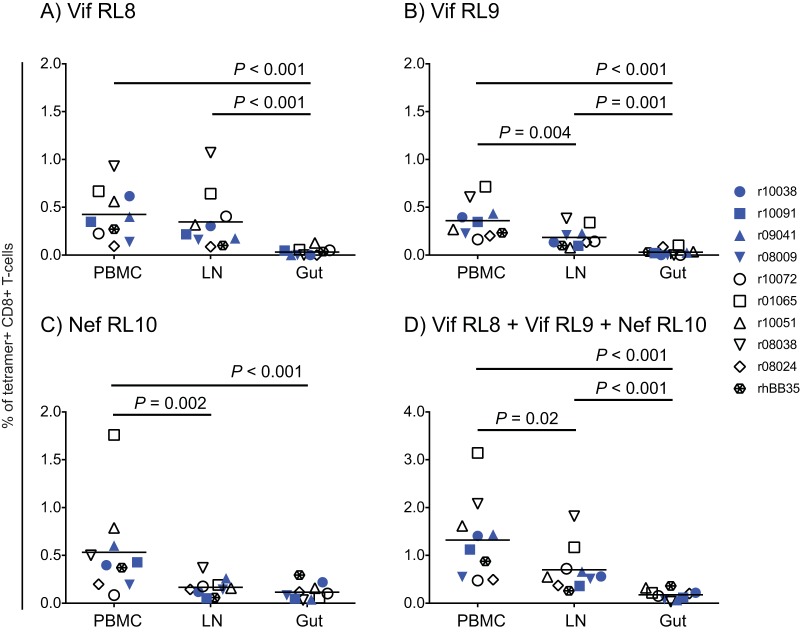
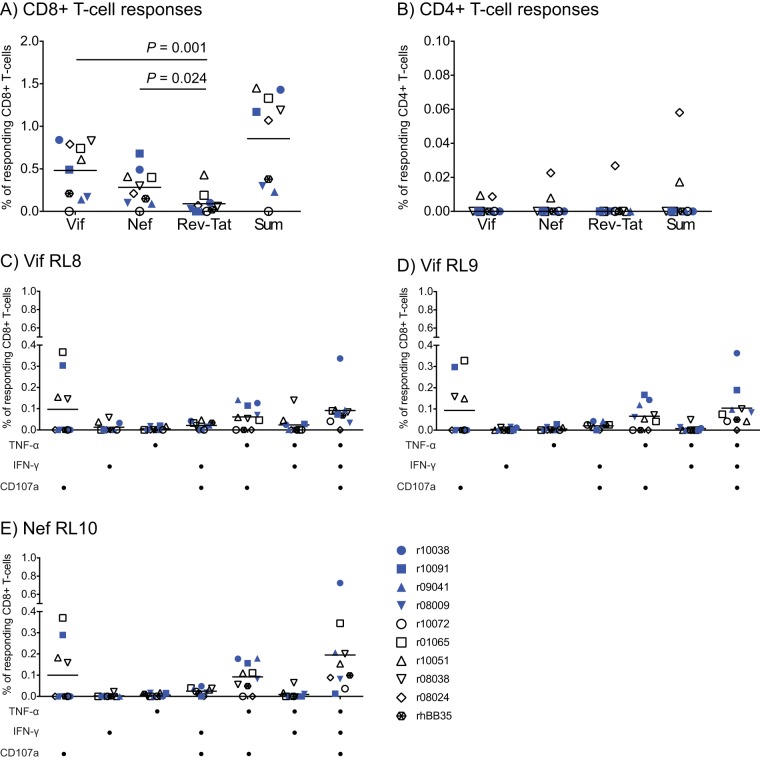
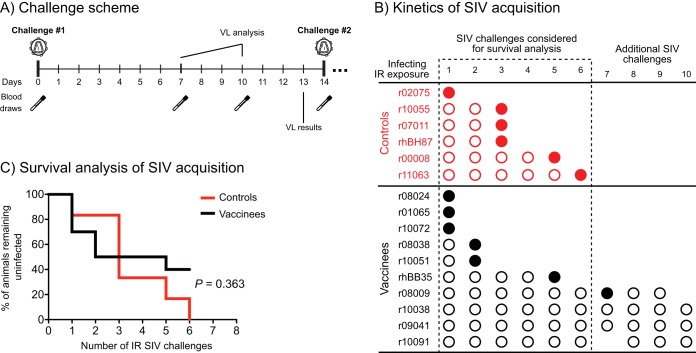
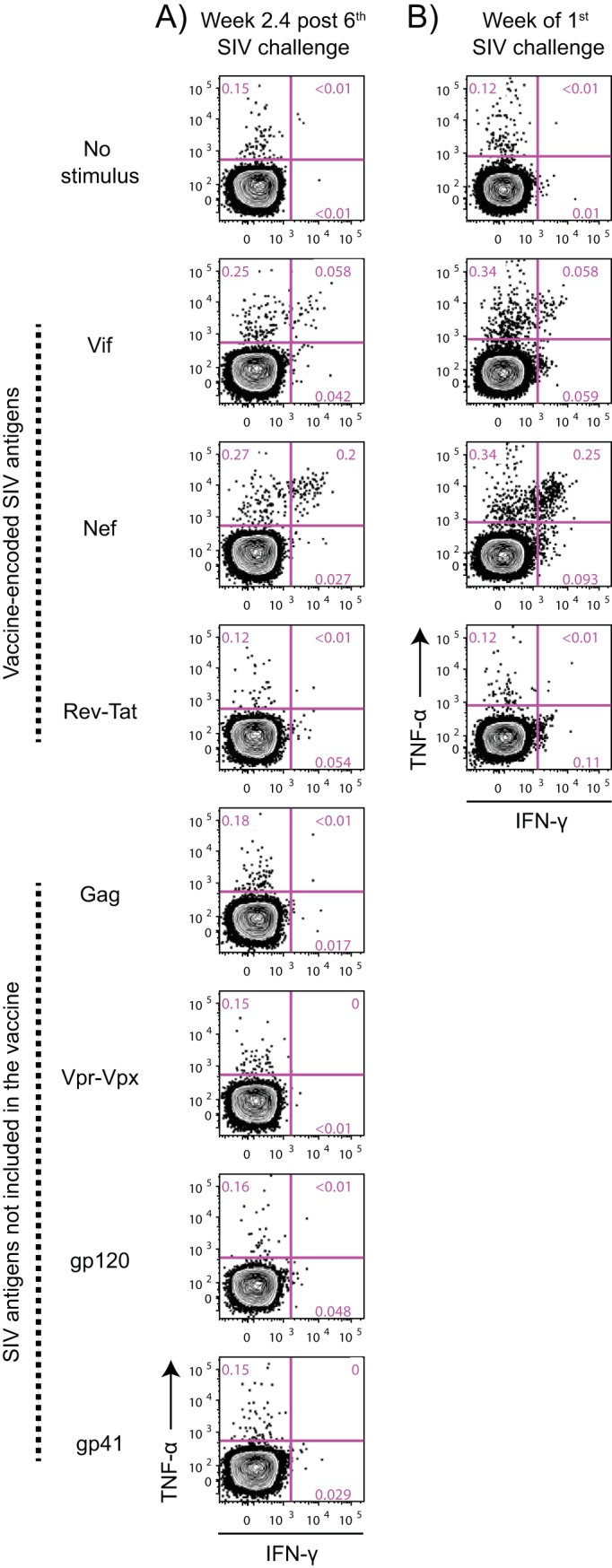
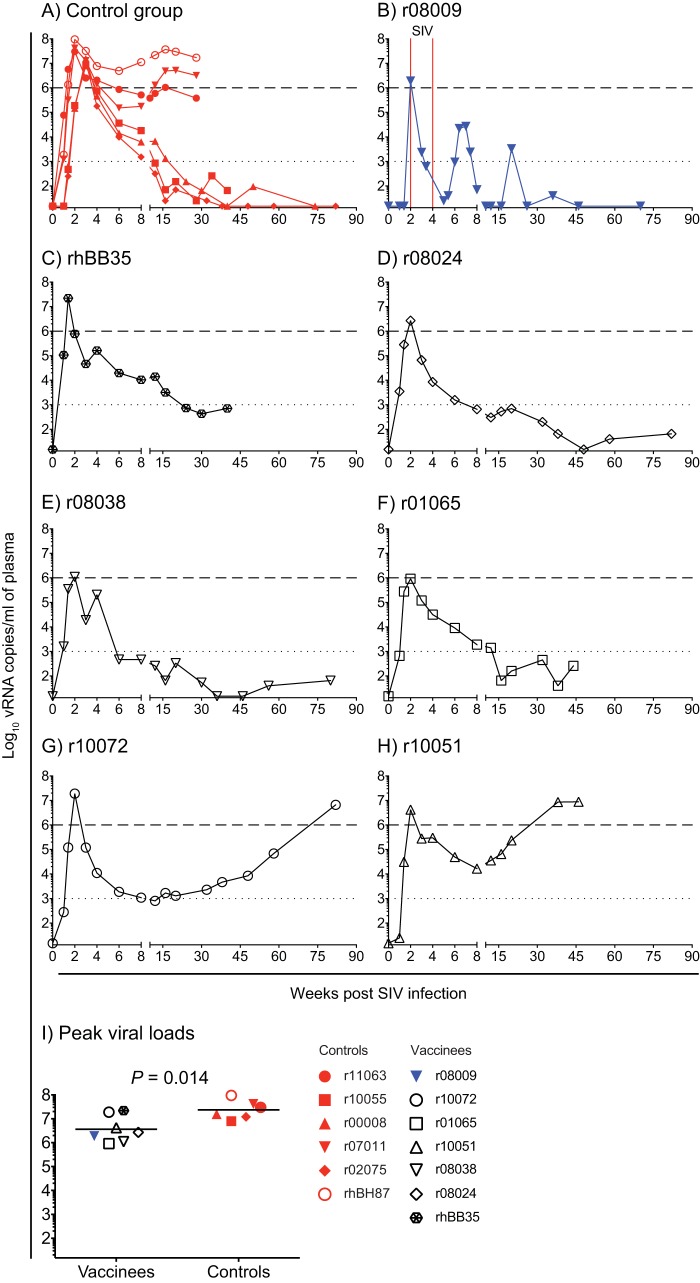
Approximately 50% of rhesus macaques (RMs) expressing the major histocompatibility complex class I (MHC-I) allele Mamu-B*08 spontaneously control chronic-phase viremia after infection with the pathogenic simian immunodeficiency virus mac239 (SIVmac239) clone. CD8+ T-cell responses in these animals are focused on immunodominant Mamu-B*08-restricted SIV epitopes in Vif and Nef, and prophylactic vaccination with these epitopes increases the incidence of elite control in SIVmac239-infected Mamu-B*08-positive (Mamu-B*08+ ) RMs. Here we evaluated if robust vaccine-elicited CD8+ T-cell responses against Vif and Nef can prevent systemic infection in Mamu-B*08+ RMs following mucosal SIV challenges. Ten Mamu-B*08+ RMs were vaccinated with a heterologous prime/boost/boost regimen encoding Vif and Nef, while six sham-vaccinated MHC-I-matched RMs served as the controls for this experiment. Vaccine-induced CD8+ T cells against Mamu-B*08-restricted SIV epitopes reached high frequencies in blood but were present at lower levels in lymph node and gut biopsy specimens. Following repeated intrarectal challenges with SIVmac239, all control RMs became infected by the sixth SIV exposure. By comparison, four vaccinees were still uninfected after six challenges, and three of them remained aviremic after 3 or 4 additional challenges. The rate of SIV acquisition in the vaccinees was numerically lower (albeit not statistically significantly) than that in the controls. However, peak viremia was significantly reduced in infected vaccinees compared to control animals. We found no T-cell markers that distinguished vaccinees that acquired SIV infection from those that did not. Additional studies will be needed to validate these findings and determine if cellular immunity can be harnessed to prevent the establishment of productive immunodeficiency virus infection.IMPORTANCE It is generally accepted that the antiviral effects of vaccine-induced classical CD8+ T-cell responses against human immunodeficiency virus (HIV) are limited to partial reductions in viremia after the establishment of productive infection. Here we show that rhesus macaques (RMs) vaccinated with Vif and Nef acquired simian immunodeficiency virus (SIV) infection at a lower (albeit not statistically significant) rate than control RMs following repeated intrarectal challenges with a pathogenic SIV clone. All animals in the present experiment expressed the elite control-associated major histocompatibility complex class I (MHC-I) molecule Mamu-B*08 that binds immunodominant epitopes in Vif and Nef. Though preliminary, these results provide tantalizing evidence that the protective efficacy of vaccine-elicited CD8+ T cells may be greater than previously thought. Future studies should examine if vaccine-induced cellular immunity can prevent systemic viral replication in RMs that do not express MHC-I alleles associated with elite control of SIV infection.
Keywords: human immunodeficiency virus; simian immunodeficiency virus; vaccines.
Copyright © 2019 American Society for Microbiology.
Figures
Similar articles
-
Acute-phase innate immune responses in SIVmac239-infected Mamu-B*08+ Indian rhesus macaques may contribute to the establishment of elite control.Front Immunol. 2024 Oct 22;15:1478063. doi: 10.3389/fimmu.2024.1478063. eCollection 2024. Front Immunol. 2024. PMID: 39502699 Free PMC article.
-
Mamu-B*17+ Rhesus Macaques Vaccinated with env, vif, and nef Manifest Early Control of SIVmac239 Replication.J Virol. 2018 Jul 31;92(16):e00690-18. doi: 10.1128/JVI.00690-18. Print 2018 Aug 15. J Virol. 2018. PMID: 29875239 Free PMC article.
-
Vaccine-Induced Simian Immunodeficiency Virus-Specific CD8+ T-Cell Responses Focused on a Single Nef Epitope Select for Escape Variants Shortly after Infection.J Virol. 2015 Nov;89(21):10802-20. doi: 10.1128/JVI.01440-15. Epub 2015 Aug 19. J Virol. 2015. PMID: 26292326 Free PMC article.
-
Antiviral CD8+ T cells in the genital tract control viral replication and delay progression to AIDS after vaginal SIV challenge in rhesus macaques immunized with virulence attenuated SHIV 89.6.J Intern Med. 2009 Jan;265(1):67-77. doi: 10.1111/j.1365-2796.2008.02051.x. J Intern Med. 2009. PMID: 19093961 Free PMC article. Review.
-
Relevance of studying T cell responses in SIV-infected rhesus macaques.Trends Microbiol. 2008 Dec;16(12):605-11. doi: 10.1016/j.tim.2008.08.010. Epub 2008 Oct 27. Trends Microbiol. 2008. PMID: 18964016 Free PMC article. Review.
Cited by
-
Acute-phase innate immune responses in SIVmac239-infected Mamu-B*08+ Indian rhesus macaques may contribute to the establishment of elite control.Front Immunol. 2024 Oct 22;15:1478063. doi: 10.3389/fimmu.2024.1478063. eCollection 2024. Front Immunol. 2024. PMID: 39502699 Free PMC article.
-
Induction of Transient Virus Replication Facilitates Antigen-Independent Isolation of SIV-Specific Monoclonal Antibodies.Mol Ther Methods Clin Dev. 2020 Feb 13;16:225-237. doi: 10.1016/j.omtm.2020.01.010. eCollection 2020 Mar 13. Mol Ther Methods Clin Dev. 2020. PMID: 32083148 Free PMC article.
-
Rhesus Cytomegalovirus-Specific CD8+ Cytotoxic T Lymphocytes Do Not Become Functionally Exhausted in Chronic SIVmac239 Infection.Front Immunol. 2020 Aug 12;11:1960. doi: 10.3389/fimmu.2020.01960. eCollection 2020. Front Immunol. 2020. PMID: 32922404 Free PMC article.
-
Recombinant Herpesvirus Vectors: Durable Immune Responses and Durable Protection against Simian Immunodeficiency Virus SIVmac239 Acquisition.J Virol. 2021 Jun 24;95(14):e0033021. doi: 10.1128/JVI.00330-21. Epub 2021 Jun 24. J Virol. 2021. PMID: 33910957 Free PMC article.
-
Env-independent protection of intrarectal SIV challenge by vaccine induction of Gag/Vif-specific CD8+ T cells but not CD4+ T cells.Mol Ther. 2022 May 4;30(5):2048-2057. doi: 10.1016/j.ymthe.2022.02.023. Epub 2022 Feb 26. Mol Ther. 2022. PMID: 35231604 Free PMC article.
References
-
- Hessell AJ, Jaworski JP, Epson E, Matsuda K, Pandey S, Kahl C, Reed J, Sutton WF, Hammond KB, Cheever TA, Barnette PT, Legasse AW, Planer S, Stanton JJ, Pegu A, Chen X, Wang K, Siess D, Burke D, Park BS, Axthelm MK, Lewis A, Hirsch VM, Graham BS, Mascola JR, Sacha JB, Haigwood NL. 2016. Early short-term treatment with neutralizing human monoclonal antibodies halts SHIV infection in infant macaques. Nat Med 22:362–368. doi:10.1038/nm.4063. - DOI - PMC - PubMed
-
- Liu J, Ghneim K, Sok D, Bosche WJ, Li Y, Chipriano E, Berkemeier B, Oswald K, Borducchi E, Cabral C, Peter L, Brinkman A, Shetty M, Jimenez J, Mondesir J, Lee B, Giglio P, Chandrashekar A, Abbink P, Colantonio A, Gittens C, Baker C, Wagner W, Lewis MG, Li W, Sekaly RP, Lifson JD, Burton DR, Barouch DH. 2016. Antibody-mediated protection against SHIV challenge includes systemic clearance of distal virus. Science 353:1045–1049. doi:10.1126/science.aag0491. - DOI - PMC - PubMed
-
- Allen TM, Altfeld M, Geer SC, Kalife ET, Moore C, O'Sullivan KM, DeSouza I, Feeney ME, Eldridge RL, Maier EL, Kaufmann DE, Lahaie MP, Reyor L, Tanzi G, Johnston MN, Brander C, Draenert R, Rockstroh JK, Jessen H, Rosenberg ES, Mallal SA, Walker BD. 2005. Selective escape from CD8+ T-cell responses represents a major driving force of human immunodeficiency virus type 1 (HIV-1) sequence diversity and reveals constraints on HIV-1 evolution. J Virol 79:13239–13249. doi:10.1128/JVI.79.21.13239-13249.2005. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
