

Functional Evolution of Subolesin/Akirin
- PMID: 30542290
- PMCID: PMC6277881
- DOI: 10.3389/fphys.2018.01612
Functional Evolution of Subolesin/Akirin
Abstract
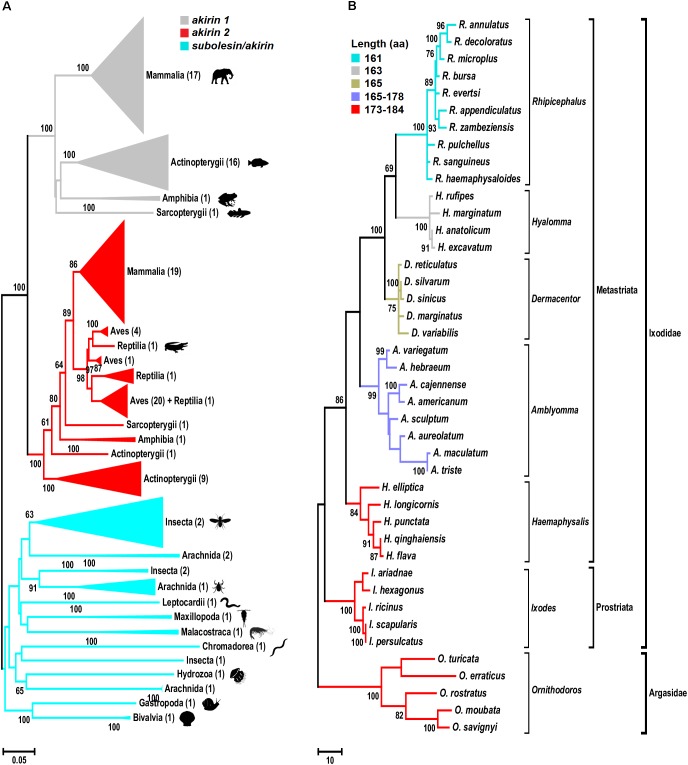
The Subolesin/Akirin constitutes a good model for the study of functional evolution because these proteins have been conserved throughout the metazoan and play a role in the regulation of different biological processes. Here, we investigated the evolutionary history of Subolesin/Akirin with recent results on their structure, protein-protein interactions and function in different species to provide insights into the functional evolution of these regulatory proteins, and their potential as vaccine antigens for the control of ectoparasite infestations and pathogen infection. The results suggest that Subolesin/Akirin evolved conserving not only its sequence and structure, but also its function and role in cell interactome and regulome in response to pathogen infection and other biological processes. This functional conservation provides a platform for further characterization of the function of these regulatory proteins, and how their evolution can meet species-specific demands. Furthermore, the conserved functional evolution of Subolesin/Akirin correlates with the protective capacity shown by these proteins in vaccine formulations for the control of different arthropod and pathogen species. These results encourage further research to characterize the structure and function of these proteins, and to develop new vaccine formulations by combining Subolesin/Akirin with interacting proteins for the control of multiple ectoparasite infestations and pathogen infection.
Keywords: Anaplasma phagocytophilum; immune response; interactome; phylogeny; regulome; tick; vaccine.
Figures









References
-
- Almazán C., Lagunes R., Villar M., Canales M., Rosario-Cruz R., Jongejan F., et al. (2010). Identification and characterization of Rhipicephalus (Boophilus) microplus candidate protective antigens for the control of cattle tick infestations. Parasitol. Res. 106 471–479. 10.1007/s00436-009-1689-1 - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources