

Gene flow mediates the role of sex chromosome meiotic drive during complex speciation
- PMID: 30543325
- PMCID: PMC6292695
- DOI: 10.7554/eLife.35468
Gene flow mediates the role of sex chromosome meiotic drive during complex speciation
Abstract
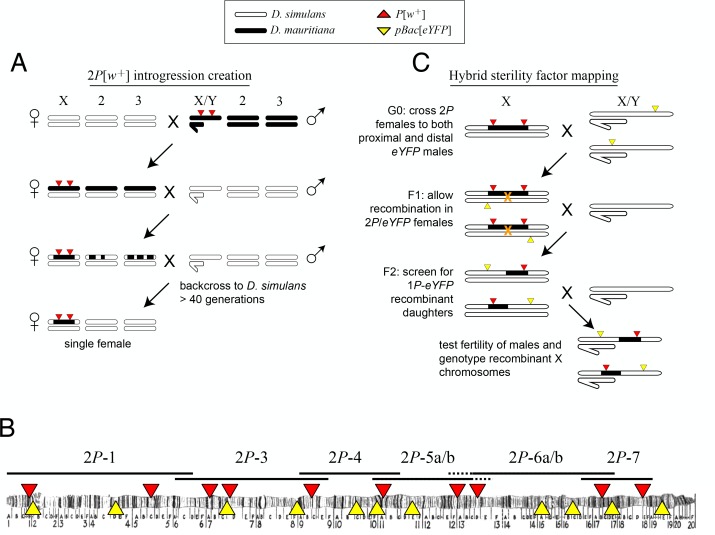
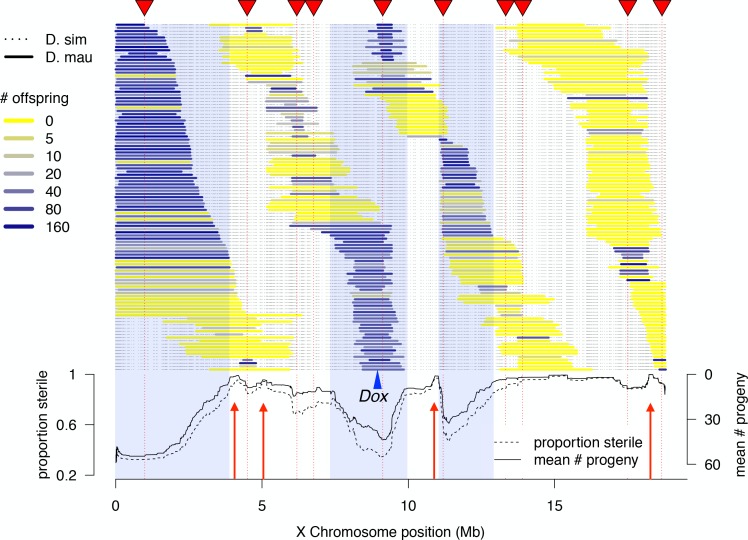
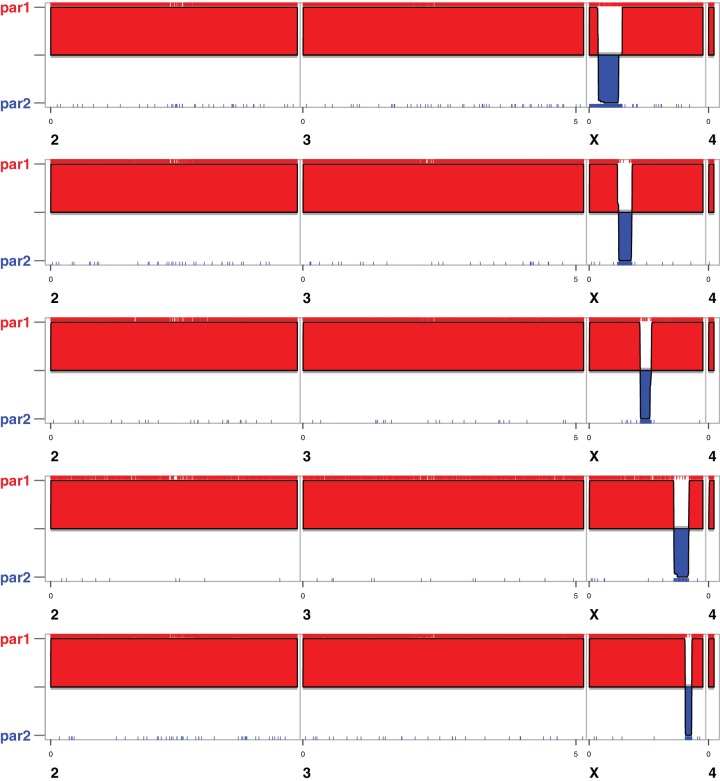
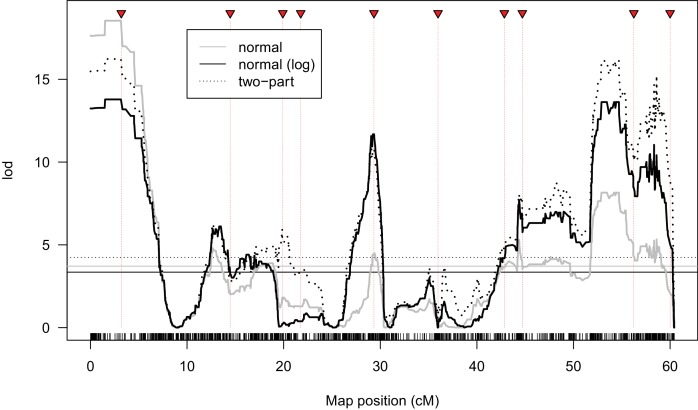
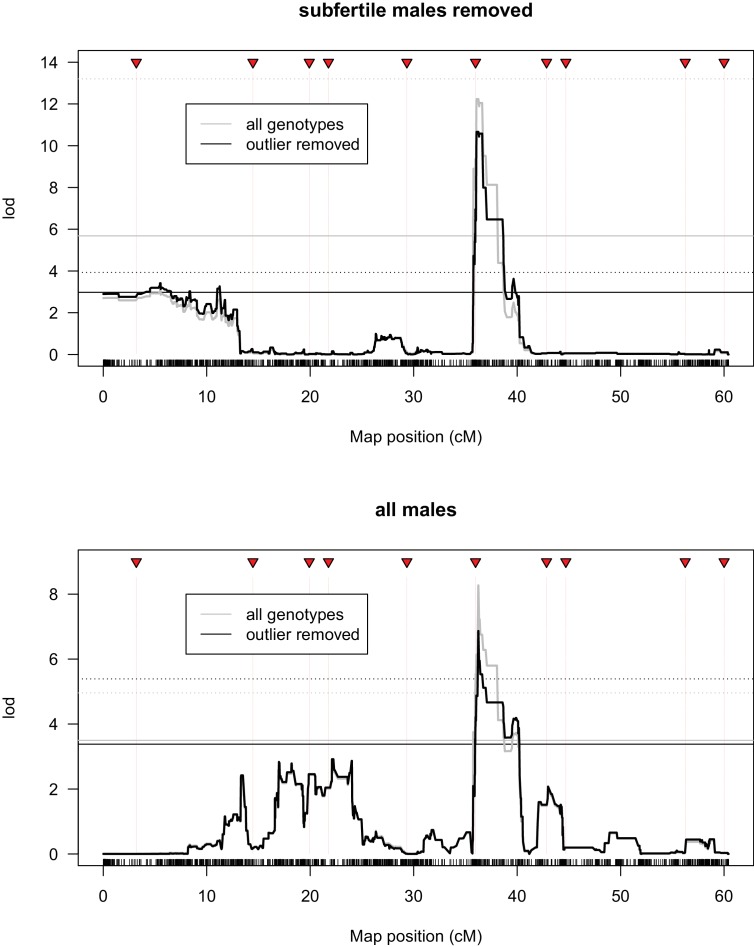
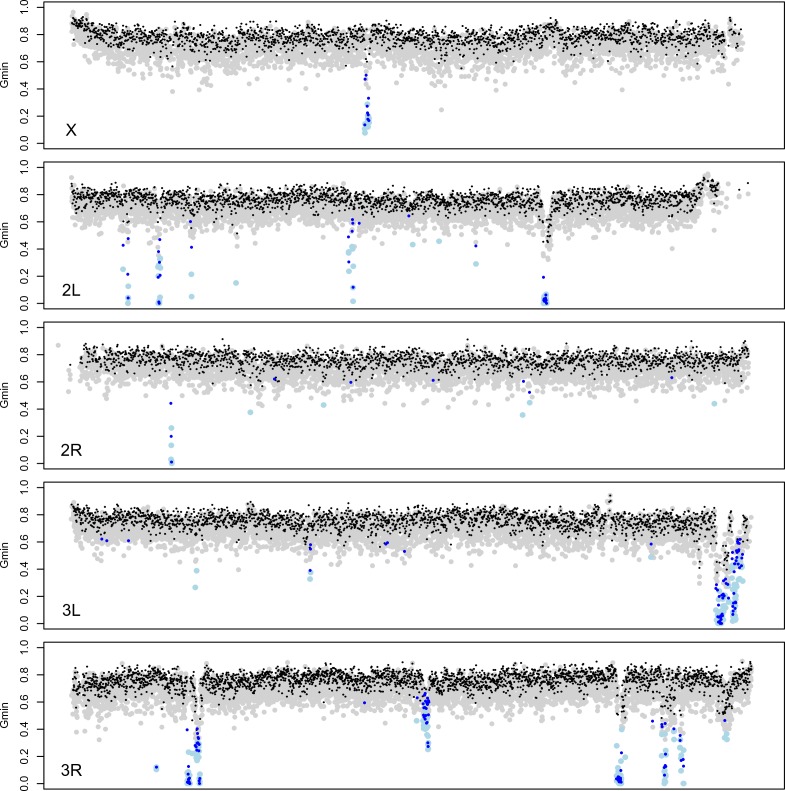
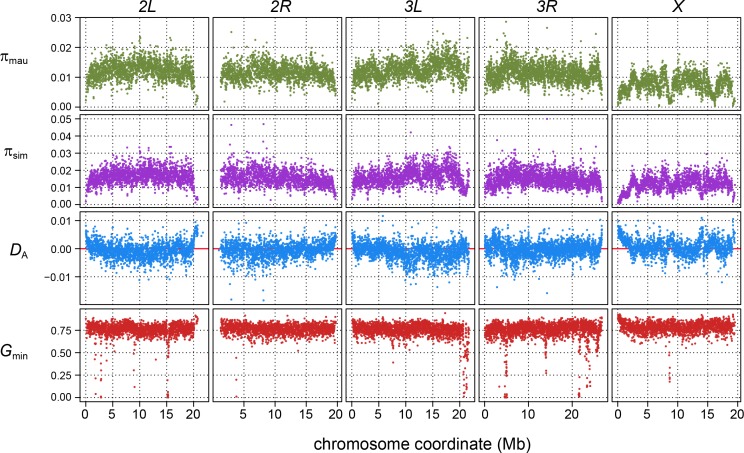
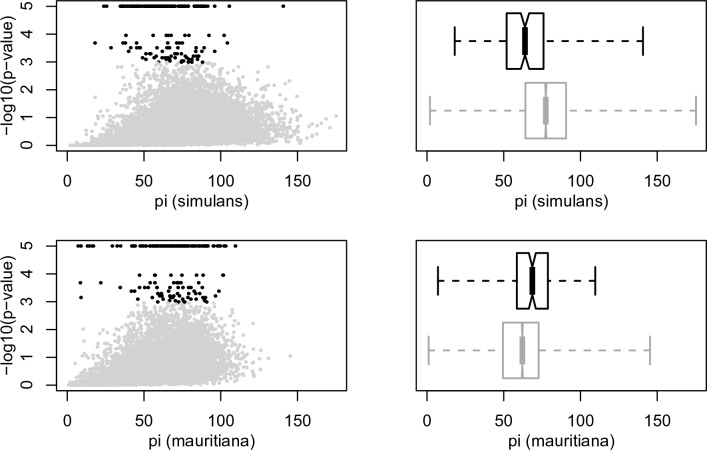
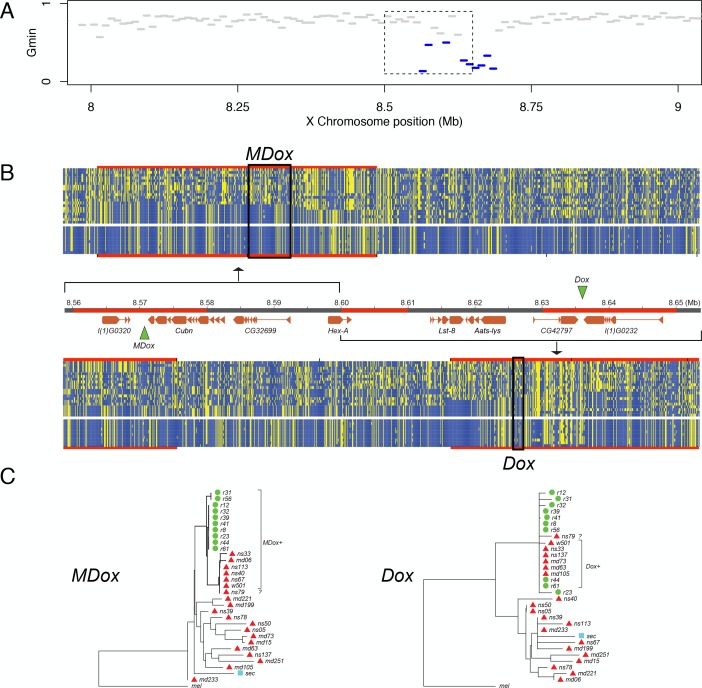
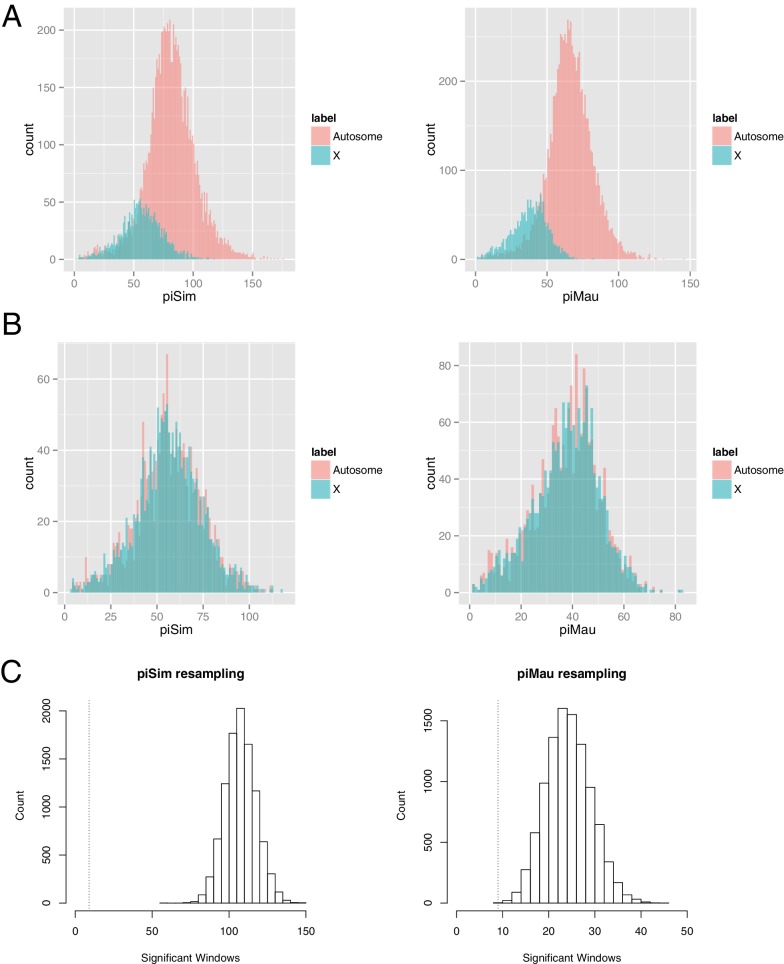
During speciation, sex chromosomes often accumulate interspecific genetic incompatibilities faster than the rest of the genome. The drive theory posits that sex chromosomes are susceptible to recurrent bouts of meiotic drive and suppression, causing the evolutionary build-up of divergent cryptic sex-linked drive systems and, incidentally, genetic incompatibilities. To assess the role of drive during speciation, we combine high-resolution genetic mapping of X-linked hybrid male sterility with population genomics analyses of divergence and recent gene flow between the fruitfly species, Drosophila mauritiana and D. simulans. Our findings reveal a high density of genetic incompatibilities and a corresponding dearth of gene flow on the X chromosome. Surprisingly, we find that a known drive element recently migrated between species and, rather than contributing to interspecific divergence, caused a strong reduction in local sequence divergence, undermining the evolution of hybrid sterility. Gene flow can therefore mediate the effects of selfish genetic elements during speciation.
Keywords: Drosophila mauritiana; Drosophila simulans; evolutionary biology; genetics; genomics; speciation.
© 2018, Meiklejohn et al.
Conflict of interest statement
CM, EL, KG, TR, SK, AG, JV, CM, DG, DS, DP No competing interests declared
Figures


Similar articles
-
Meiotic Drive and Speciation.Annu Rev Genet. 2024 Nov;58(1):341-363. doi: 10.1146/annurev-genet-111523-102603. Annu Rev Genet. 2024. PMID: 39585909 Review.
-
Genome diversity and divergence in Drosophila mauritiana: multiple signatures of faster X evolution.Genome Biol Evol. 2014 Sep 4;6(9):2444-58. doi: 10.1093/gbe/evu198. Genome Biol Evol. 2014. PMID: 25193308 Free PMC article.
-
Genetic dissection of hybrid incompatibilities between Drosophila simulans and D. mauritiana. I. Differential accumulation of hybrid male sterility effects on the X and autosomes.Genetics. 2003 Aug;164(4):1383-97. doi: 10.1093/genetics/164.4.1383. Genetics. 2003. PMID: 12930747 Free PMC article.
-
Interspecific Y chromosome variation is sufficient to rescue hybrid male sterility and is influenced by the grandparental origin of the chromosomes.Heredity (Edinb). 2016 Jun;116(6):516-22. doi: 10.1038/hdy.2016.11. Epub 2016 Mar 16. Heredity (Edinb). 2016. PMID: 26980343 Free PMC article.
-
Selfish X chromosomes and speciation.Mol Ecol. 2018 Oct;27(19):3772-3782. doi: 10.1111/mec.14471. Epub 2018 Jan 29. Mol Ecol. 2018. PMID: 29281152 Review.
Cited by
-
Widespread introgression across a phylogeny of 155 Drosophila genomes.Curr Biol. 2022 Jan 10;32(1):111-123.e5. doi: 10.1016/j.cub.2021.10.052. Epub 2021 Nov 16. Curr Biol. 2022. PMID: 34788634 Free PMC article.
-
Patterns and mechanisms of sex ratio distortion in the Collaborative Cross mouse mapping population.Genetics. 2021 Nov 5;219(3):iyab136. doi: 10.1093/genetics/iyab136. Genetics. 2021. PMID: 34740238 Free PMC article.
-
The Y Chromosome as a Battleground for Intragenomic Conflict.Trends Genet. 2020 Jul;36(7):510-522. doi: 10.1016/j.tig.2020.04.008. Epub 2020 May 21. Trends Genet. 2020. PMID: 32448494 Free PMC article. Review.
-
The Evolutionary History of Drosophila simulans Y Chromosomes Reveals Molecular Signatures of Resistance to Sex Ratio Meiotic Drive.Mol Biol Evol. 2023 Jul 5;40(7):msad152. doi: 10.1093/molbev/msad152. Mol Biol Evol. 2023. PMID: 37401458 Free PMC article.
-
Satellite DNA-mediated diversification of a sex-ratio meiotic drive gene family in Drosophila.Nat Ecol Evol. 2021 Dec;5(12):1604-1612. doi: 10.1038/s41559-021-01543-8. Epub 2021 Sep 6. Nat Ecol Evol. 2021. PMID: 34489561 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
