

Modulation of Innate Immune Responses by the Influenza A NS1 and PA-X Proteins
- PMID: 30545063
- PMCID: PMC6315843
- DOI: 10.3390/v10120708
Modulation of Innate Immune Responses by the Influenza A NS1 and PA-X Proteins
Abstract
Influenza A viruses (IAV) can infect a broad range of animal hosts, including humans. In humans, IAV causes seasonal annual epidemics and occasional pandemics, representing a serious public health and economic problem, which is most effectively prevented through vaccination. The defense mechanisms that the host innate immune system provides restrict IAV replication and infection. Consequently, to successfully replicate in interferon (IFN)-competent systems, IAV has to counteract host antiviral activities, mainly the production of IFN and the activities of IFN-induced host proteins that inhibit virus replication. The IAV multifunctional proteins PA-X and NS1 are virulence factors that modulate the innate immune response and virus pathogenicity. Notably, these two viral proteins have synergistic effects in the inhibition of host protein synthesis in infected cells, although using different mechanisms of action. Moreover, the control of innate immune responses by the IAV NS1 and PA-X proteins is subject to a balance that can determine virus pathogenesis and fitness, and recent evidence shows co-evolution of these proteins in seasonal viruses, indicating that they should be monitored for enhanced virulence. Importantly, inhibition of host gene expression by the influenza NS1 and/or PA-X proteins could be explored to develop improved live-attenuated influenza vaccines (LAIV) by modulating the ability of the virus to counteract antiviral host responses. Likewise, both viral proteins represent a reasonable target for the development of new antivirals for the control of IAV infections. In this review, we summarize the role of IAV NS1 and PA-X in controlling the antiviral response during viral infection.
Keywords: NS1; PA; PA-X; cytokines; host antiviral response; influenza a virus; innate immunity; interferon; pathogenesis; vaccine; virus evolution; virus–host interactions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
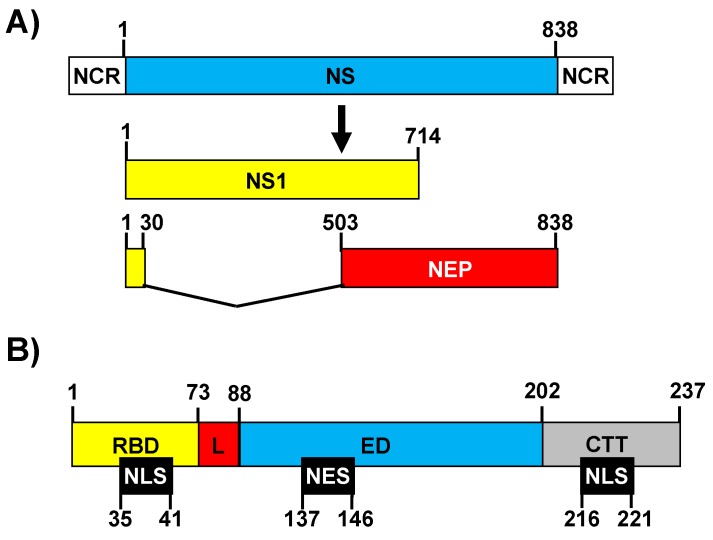
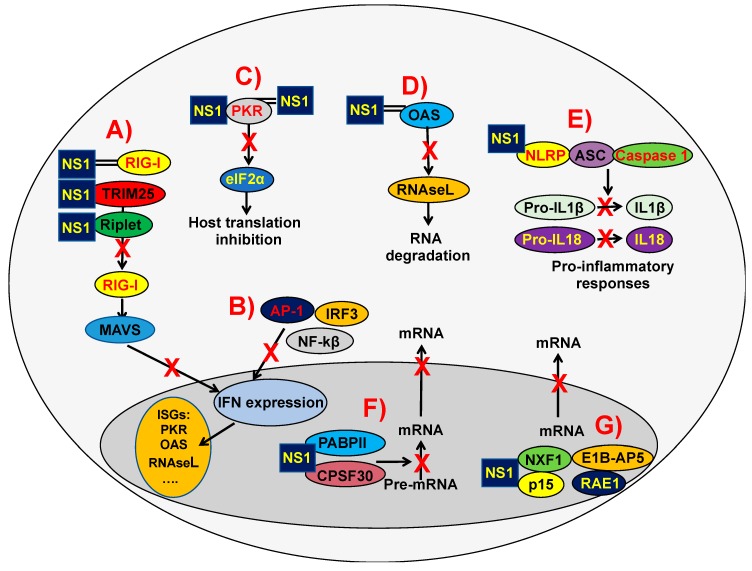
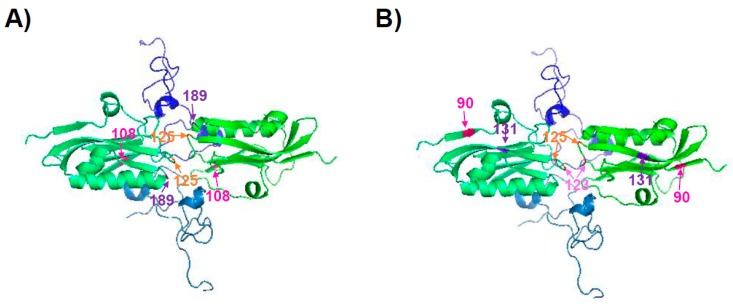
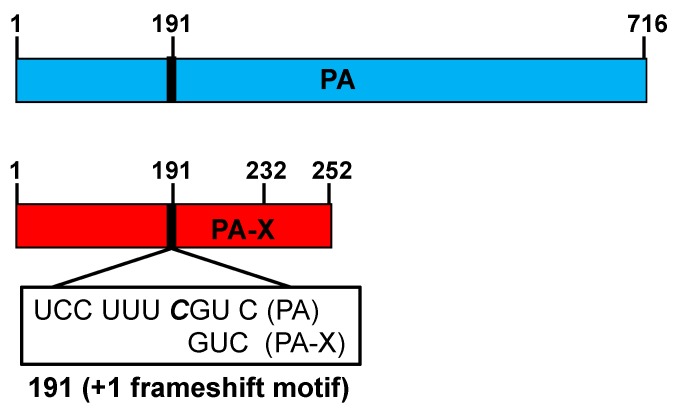
References
-
- Palese P., Shaw M.L. Orthomyxoviridae: The viruses and their replication. In: Knipe D., Howley P., editors. Fields Virology. Lippincott Williams & Wilkins; Philadelphia, PA, USA: 2006. pp. 1648–1689.
-
- Bailey E.S., Choi J.Y., Fieldhouse J.K., Borkenhagen L.K., Zemke J., Zhang D., Gray G.C. The continual threat of influenza virus infections at the human-animal interface: What is new from a one health perspective? Evol. Med. Public Health. 2018;2018:192–198. doi: 10.1093/emph/eoy013. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
