

Integrative functional genomic analysis of human brain development and neuropsychiatric risks
- PMID: 30545854
- PMCID: PMC6413317
- DOI: 10.1126/science.aat7615
Integrative functional genomic analysis of human brain development and neuropsychiatric risks
Abstract
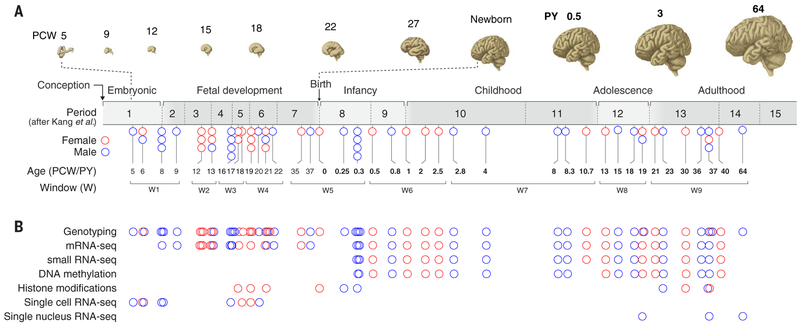
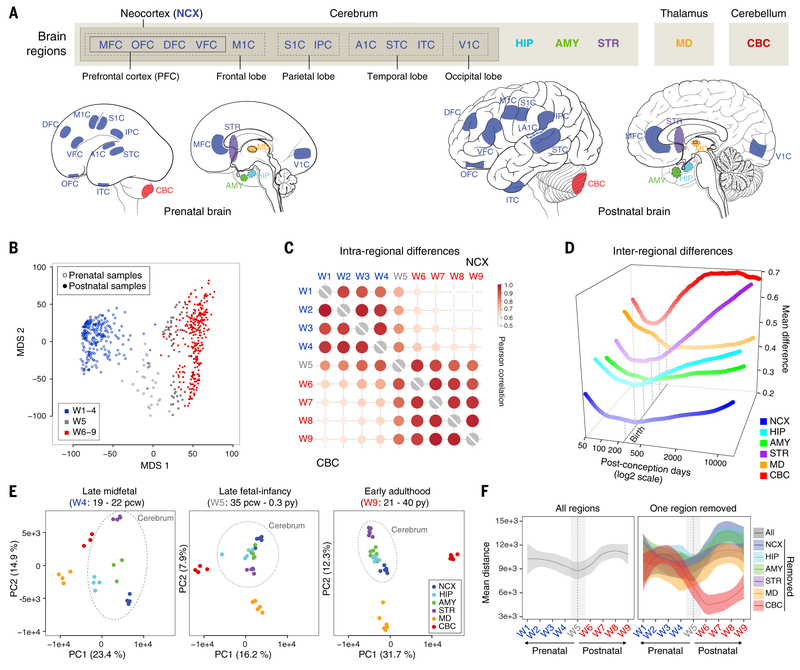
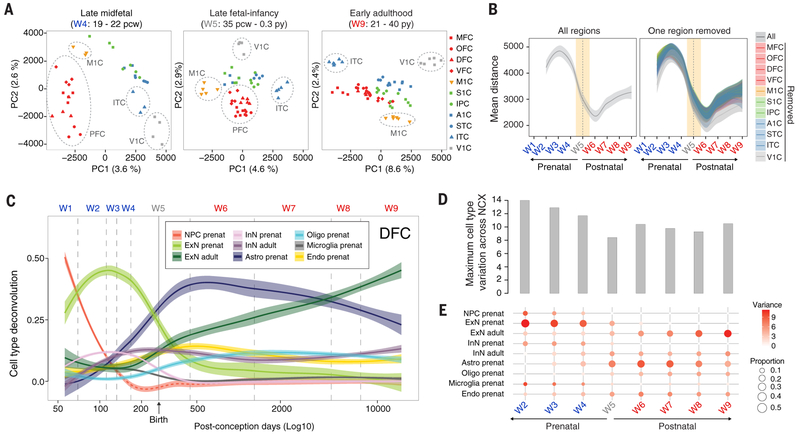
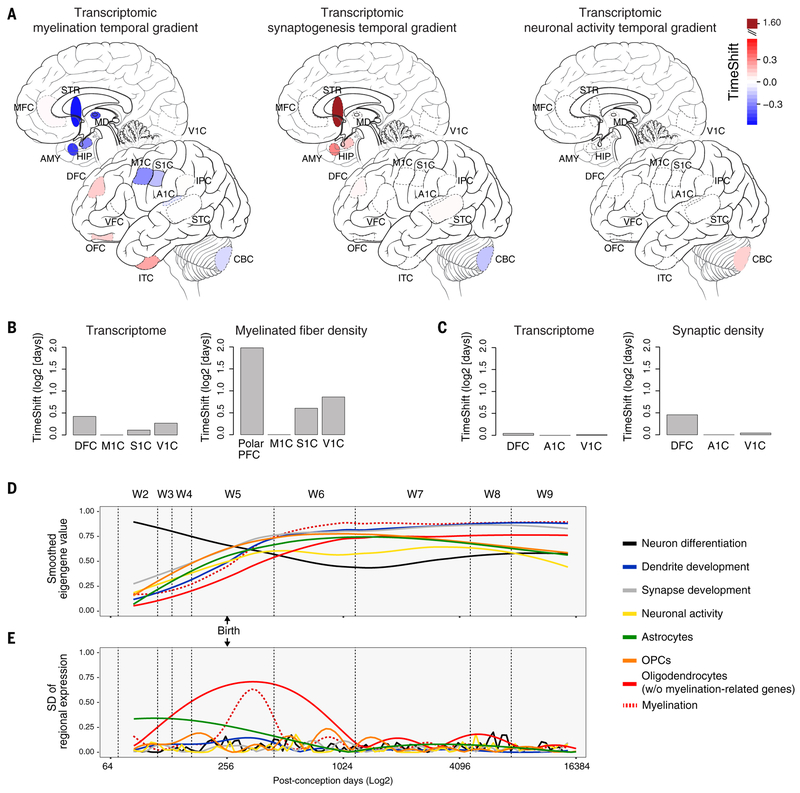
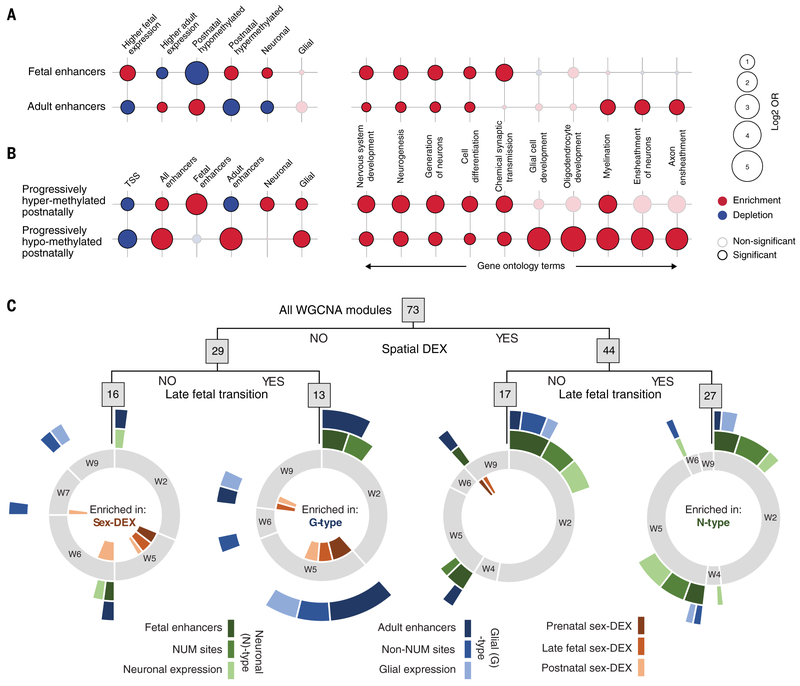
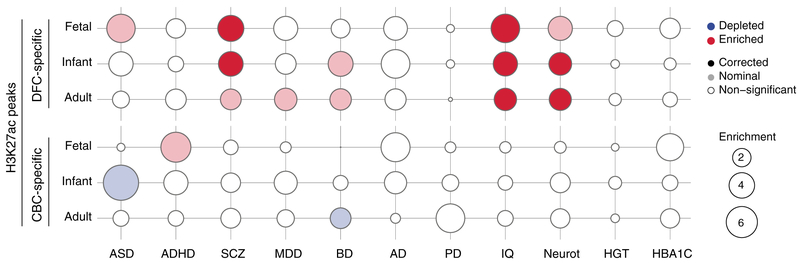
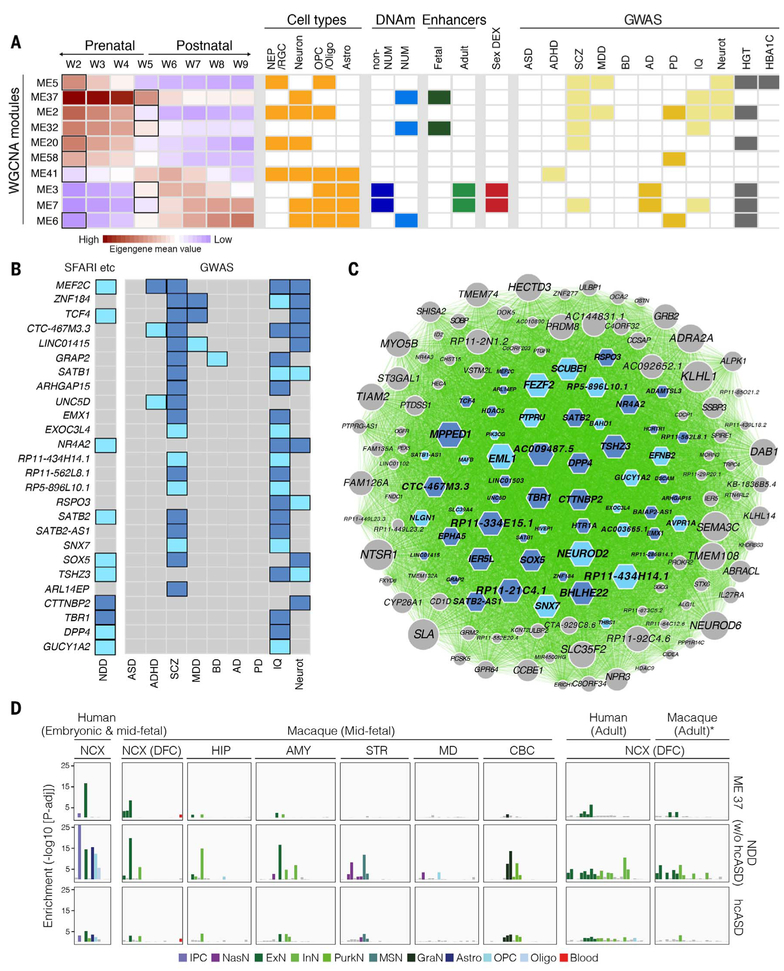
To broaden our understanding of human neurodevelopment, we profiled transcriptomic and epigenomic landscapes across brain regions and/or cell types for the entire span of prenatal and postnatal development. Integrative analysis revealed temporal, regional, sex, and cell type-specific dynamics. We observed a global transcriptomic cup-shaped pattern, characterized by a late fetal transition associated with sharply decreased regional differences and changes in cellular composition and maturation, followed by a reversal in childhood-adolescence, and accompanied by epigenomic reorganizations. Analysis of gene coexpression modules revealed relationships with epigenomic regulation and neurodevelopmental processes. Genes with genetic associations to brain-based traits and neuropsychiatric disorders (including MEF2C, SATB2, SOX5, TCF4, and TSHZ3) converged in a small number of modules and distinct cell types, revealing insights into neurodevelopment and the genomic basis of neuropsychiatric risks.
Copyright © 2018 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Grants and funding
- R01 MH110928/MH/NIMH NIH HHS/United States
- R01 MH094714/MH/NIMH NIH HHS/United States
- U01 MH103365/MH/NIMH NIH HHS/United States
- R01 MH110905/MH/NIMH NIH HHS/United States
- R00 MH113823/MH/NIMH NIH HHS/United States
- R01 MH110927/MH/NIMH NIH HHS/United States
- R21 MH109956/MH/NIMH NIH HHS/United States
- S10 OD018521/OD/NIH HHS/United States
- U01 MH103392/MH/NIMH NIH HHS/United States
- R01 MH109677/MH/NIMH NIH HHS/United States
- U01 MH103346/MH/NIMH NIH HHS/United States
- RC2 MH089921/MH/NIMH NIH HHS/United States
- R01 MH109901/MH/NIMH NIH HHS/United States
- U01 MH103339/MH/NIMH NIH HHS/United States
- G0700089/MRC_/Medical Research Council/United Kingdom
- R01 MH110926/MH/NIMH NIH HHS/United States
- P50 MH106934/MH/NIMH NIH HHS/United States
- I01 BX002876/BX/BLRD VA/United States
- R21 MH103877/MH/NIMH NIH HHS/United States
- RC2 MH090047/MH/NIMH NIH HHS/United States
- R01 DA043247/DA/NIDA NIH HHS/United States
- EP-C-14-005/EPA/EPA/United States
- R01 MH111721/MH/NIMH NIH HHS/United States
- U01 MH103340/MH/NIMH NIH HHS/United States
- R01 MH110920/MH/NIMH NIH HHS/United States
- R24 HD000836/HD/NICHD NIH HHS/United States
- R01 HG009658/HG/NHGRI NIH HHS/United States
- R21 MH102791/MH/NIMH NIH HHS/United States
- R01 MH105898/MH/NIMH NIH HHS/United States
- R21 MH105881/MH/NIMH NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- K01 MH109772/MH/NIMH NIH HHS/United States
- S10 OD030363/OD/NIH HHS/United States
- S10 OD018164/OD/NIH HHS/United States
- R01 MH110921/MH/NIMH NIH HHS/United States
- RC2 MH089929/MH/NIMH NIH HHS/United States
- MR/L010305/1/MRC_/Medical Research Council/United Kingdom
- R21 MH105853/MH/NIMH NIH HHS/United States
- R01 MH109715/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
