

Toll-like receptor 3 activation increases voluntary alcohol intake in C57BL/6J male mice
- PMID: 30550931
- PMCID: PMC6399060
- DOI: 10.1016/j.bbi.2018.12.004
Toll-like receptor 3 activation increases voluntary alcohol intake in C57BL/6J male mice
Abstract
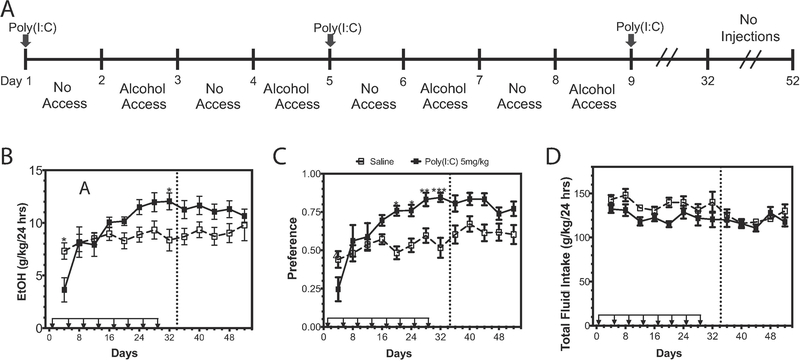
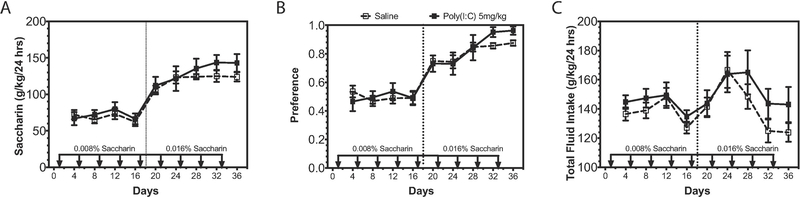
Many genes differentially expressed in brain tissue from human alcoholics and animals that have consumed large amounts of alcohol are components of the innate immune toll-like receptor (TLR) pathway. TLRs initiate inflammatory responses via two branches: (1) MyD88-dependent or (2) TRIF-dependent. All TLRs signal through MyD88 except TLR3. Prior work demonstrated a direct role for MyD88-dependent signaling in regulation of alcohol consumption. However, the role of TLR3 as a potential regulator of excessive alcohol drinking has not previously been investigated. To test the possibility TLR3 activation regulates alcohol consumption, we injected mice with the TLR3 agonist polyinosinic:polycytidylic acid (poly(I:C)) and tested alcohol consumption in an every-other-day two-bottle choice test. Poly(I:C) produced a persistent increase in alcohol intake that developed over several days. Repeated poly(I:C) and ethanol exposure altered innate immune transcript abundance; increased levels of TRIF-dependent pathway components correlated with increased alcohol consumption. Administration of poly(I:C) before exposure to alcohol did not alter alcohol intake, suggesting that poly(I:C) and ethanol must be present together to change drinking behavior. To determine which branch of TLR signaling mediates poly(I:C)-induced changes in drinking behavior, we tested either mice lacking MyD88 or mice administered a TLR3/dsRNA complex inhibitor. MyD88 null mutants showed poly(I:C)-induced increases in alcohol intake. In contrast, mice pretreated with a TLR3/dsRNA complex inhibitor reduced their alcohol intake, suggesting poly(I:C)-induced escalations in alcohol intake are, at least partially, dependent on TLR3. Together, these results strongly suggest that TLR3-dependent signaling drives excessive alcohol drinking behavior.
Keywords: Alcohol use disorder; Cytokines; Drinking; Ethanol; Neuroimmune; Poly(I:C); Toll-like receptors.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors report no biomedical financial interests or potential conflicts of interest.
Figures






References
-
- Montesinos J, Alfonso-Loeches S, Guerri C (2016): Impact of the Innate Immune Response in the Actions of Ethanol on the Central Nervous System. Alcohol Clin Exp Res 40:2260–2270. - PubMed
-
- Liu J, Lewohl JM, Harris RA, Iyer VR, Dodd PR, Randall PK, et al. (2006): Patterns of gene expression in the frontal cortex discriminate alcoholic from nonalcoholic individuals. Neuropsychopharmacology 31:1574–1582. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
