

Salmonella Persist in Activated Macrophages in T Cell-Sparse Granulomas but Are Contained by Surrounding CXCR3 Ligand-Positioned Th1 Cells
- PMID: 30552021
- PMCID: PMC6301113
- DOI: 10.1016/j.immuni.2018.10.009
Salmonella Persist in Activated Macrophages in T Cell-Sparse Granulomas but Are Contained by Surrounding CXCR3 Ligand-Positioned Th1 Cells
Abstract
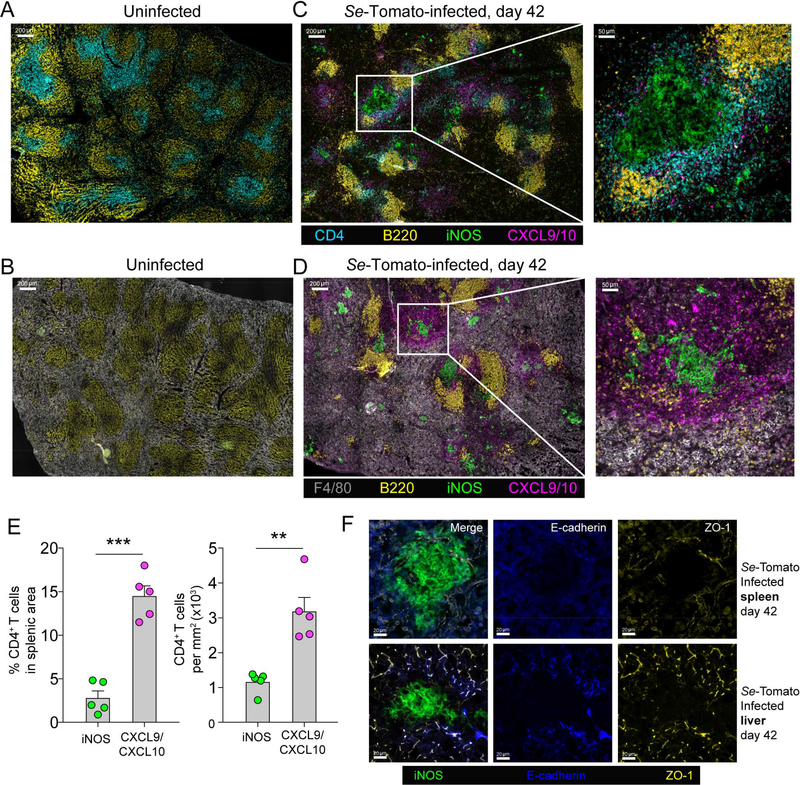
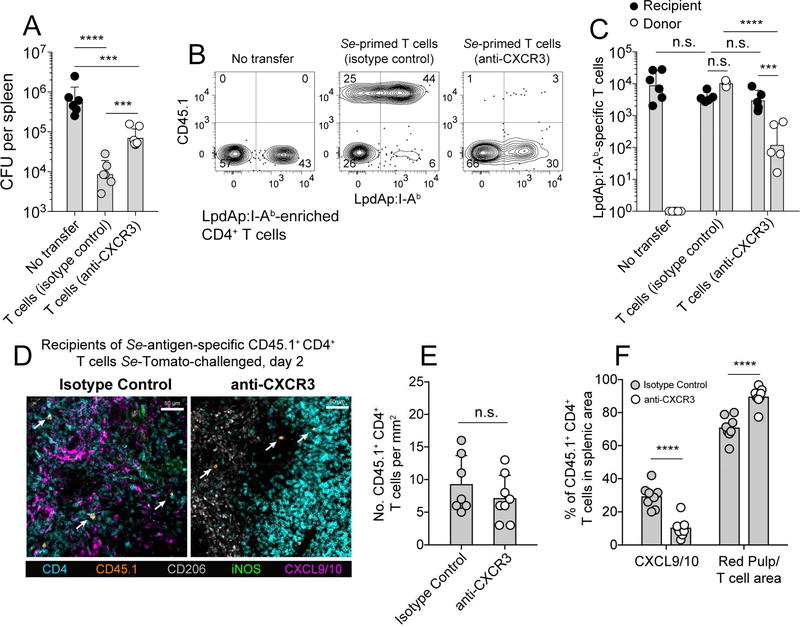
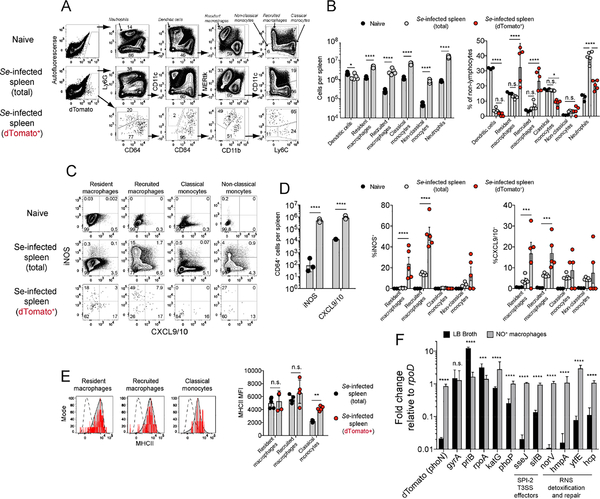
Salmonella enterica (Se) bacteria cause persistent intracellular infections while stimulating a robust interferon-γ-producing CD4+ T (Th1) cell response. We addressed this paradox of concomitant infection and immunity by tracking fluorescent Se organisms in mice. Se bacteria persisted in nitric oxide synthase (iNOS)-producing resident and recruited macrophages while inducing genes related to protection from nitric oxide. Se-infected cells occupied iNOS+ splenic granulomas that excluded T cells but were surrounded by mononuclear phagocytes producing the chemokines CXCL9 and CXCL10, and Se epitope-specific Th1 cells expressing CXCR3, the receptor for these chemokines. Blockade of CXCR3 inhibited Th1 occupancy of CXCL9/10-dense regions, reduced activation of the Th1 cells, and led to increased Se growth. Thus, intracellular Se bacteria survive in their hosts by counteracting toxic products of the innate immune response and by residing in T cell-sparse granulomas, away from abundant Th1 cells positioned via CXCR3 in a bordering region that act to limit infection.
Keywords: CXCR3; Th1; granuloma; phagosomal pathogen.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures




Similar articles
-
Blockade of Th1 chemokine receptors ameliorates pulmonary granulomatosis in mice.Eur Respir J. 2011 Aug;38(2):415-24. doi: 10.1183/09031936.00070610. Epub 2011 Jan 27. Eur Respir J. 2011. PMID: 21273392
-
Liver inflammation in a mouse model of Th1 hepatitis despite the absence of invariant NKT cells or the Th1 chemokine receptors CXCR3 and CCR5.Lab Invest. 2012 Oct;92(10):1461-71. doi: 10.1038/labinvest.2012.104. Epub 2012 Aug 20. Lab Invest. 2012. PMID: 22906987 Free PMC article.
-
CXCR3 ligands contribute to Th1-induced inflammation but not to homing of Th1 cells into the lung.Exp Lung Res. 2008 Sep;34(7):391-407. doi: 10.1080/01902140802221987. Exp Lung Res. 2008. PMID: 18716926 Free PMC article.
-
CXCR3 Ligands in Cancer and Autoimmunity, Chemoattraction of Effector T Cells, and Beyond.Front Immunol. 2020 May 29;11:976. doi: 10.3389/fimmu.2020.00976. eCollection 2020. Front Immunol. 2020. PMID: 32547545 Free PMC article. Review.
-
Hypersensitivity pneumonitis and alpha-chemokines.Clin Ter. 2017 Mar-Apr;168(2):e140-e145. doi: 10.7417/CT.2017.1996. Clin Ter. 2017. PMID: 28383627 Review.
Cited by
-
Failure of CD4 T Cell-Deficient Hosts To Control Chronic Nontyphoidal Salmonella Infection Leads to Exacerbated Inflammation, Chronic Anemia, and Altered Myelopoiesis.Infect Immun. 2020 Dec 15;89(1):e00417-20. doi: 10.1128/IAI.00417-20. Print 2020 Dec 15. Infect Immun. 2020. PMID: 33046510 Free PMC article.
-
Designing a novel mRNA vaccine against Vibrio harveyi infection in fish: an immunoinformatics approach.Genomics Inform. 2022 Mar;20(1):e11. doi: 10.5808/gi.21065. Epub 2022 Mar 31. Genomics Inform. 2022. PMID: 35399010 Free PMC article.
-
JAK inhibition in a patient with a STAT1 gain-of-function variant reveals STAT1 dysregulation as a common feature of aplastic anemia.Med. 2022 Jan 14;3(1):42-57.e5. doi: 10.1016/j.medj.2021.12.003. Med. 2022. PMID: 35590143 Free PMC article.
-
Hacking the host: exploitation of macrophage polarization by intracellular bacterial pathogens.Pathog Dis. 2020 Feb 1;78(1):ftaa009. doi: 10.1093/femspd/ftaa009. Pathog Dis. 2020. PMID: 32068828 Free PMC article. Review.
-
A novel strategy for developing vaccine candidate against Jaagsiekte sheep retrovirus from the envelope and gag proteins: an in-silico approach.BMC Vet Res. 2022 Sep 10;18(1):343. doi: 10.1186/s12917-022-03431-0. BMC Vet Res. 2022. PMID: 36085036 Free PMC article.
References
-
- Belkaid Y, Piccirillo CA, Mendez S, Shevach EM, and Sacks DL (2002). CD4+CD25+ regulatory T cells control Leishmania major persistence and immunity. Nature 420, 502–507. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
