

RSC-Associated Subnucleosomes Define MNase-Sensitive Promoters in Yeast
- PMID: 30554944
- PMCID: PMC6475595
- DOI: 10.1016/j.molcel.2018.10.046
RSC-Associated Subnucleosomes Define MNase-Sensitive Promoters in Yeast
Abstract
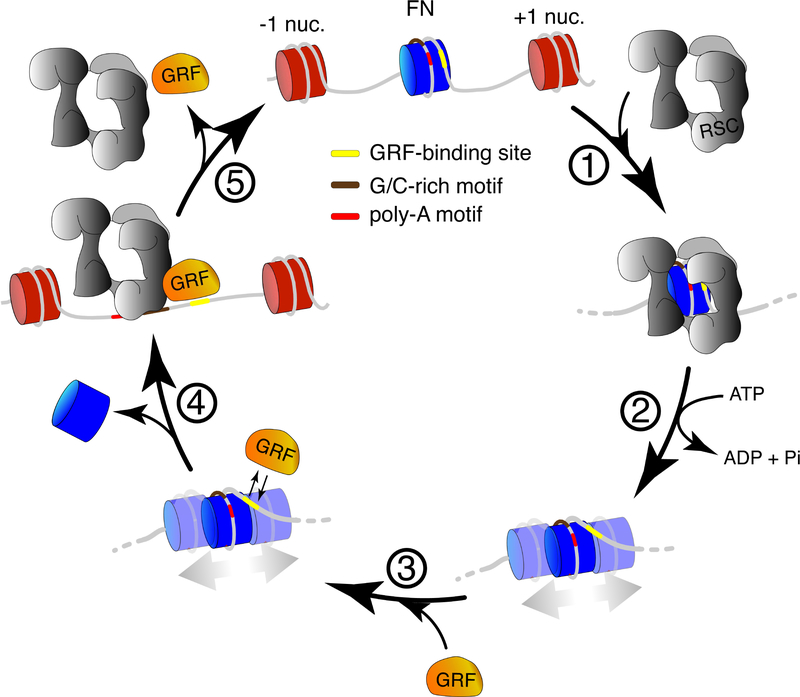
The classic view of nucleosome organization at active promoters is that two well-positioned nucleosomes flank a nucleosome-depleted region (NDR). However, this view has been recently disputed by contradictory reports as to whether wider (≳150 bp) NDRs instead contain unstable, micrococcal nuclease-sensitive ("fragile") nucleosomal particles. To determine the composition of fragile particles, we introduce CUT&RUN.ChIP, in which targeted nuclease cleavage and release is followed by chromatin immunoprecipitation. We find that fragile particles represent the occupancy of the RSC (remodeling the structure of chromatin) nucleosome remodeling complex and RSC-bound, partially unwrapped nucleosomal intermediates. We also find that general regulatory factors (GRFs) bind to partially unwrapped nucleosomes at these promoters. We propose that RSC binding and its action cause nucleosomes to unravel, facilitate subsequent binding of GRFs, and constitute a dynamic cycle of nucleosome deposition and clearance at the subset of wide Pol II promoter NDRs.
Keywords: CUT&RUN; fragile nucleosome; general regulatory factor; nucleosome dynamics; nucleosome-depleted region.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interest
The authors declare no competing interests.
Figures
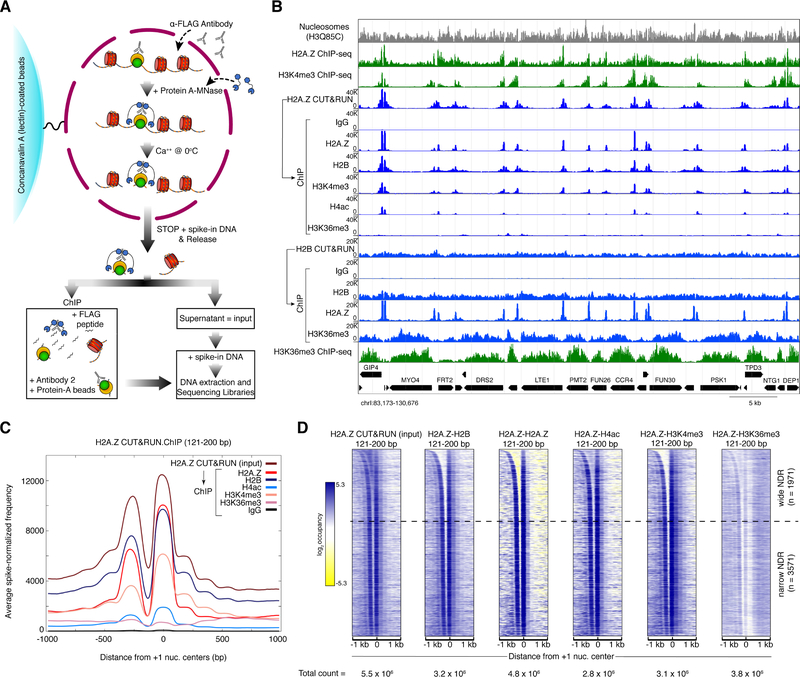
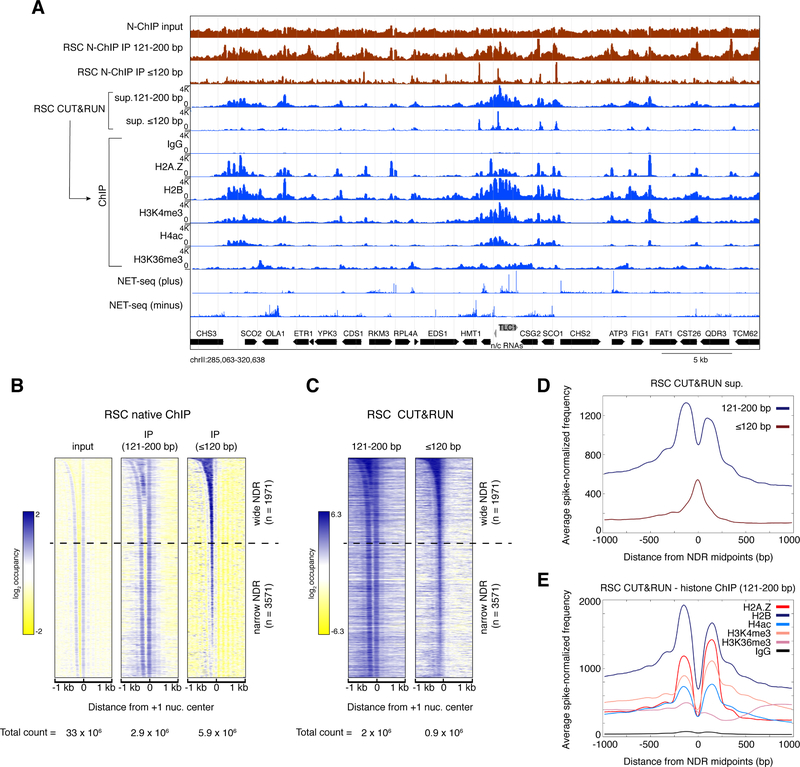
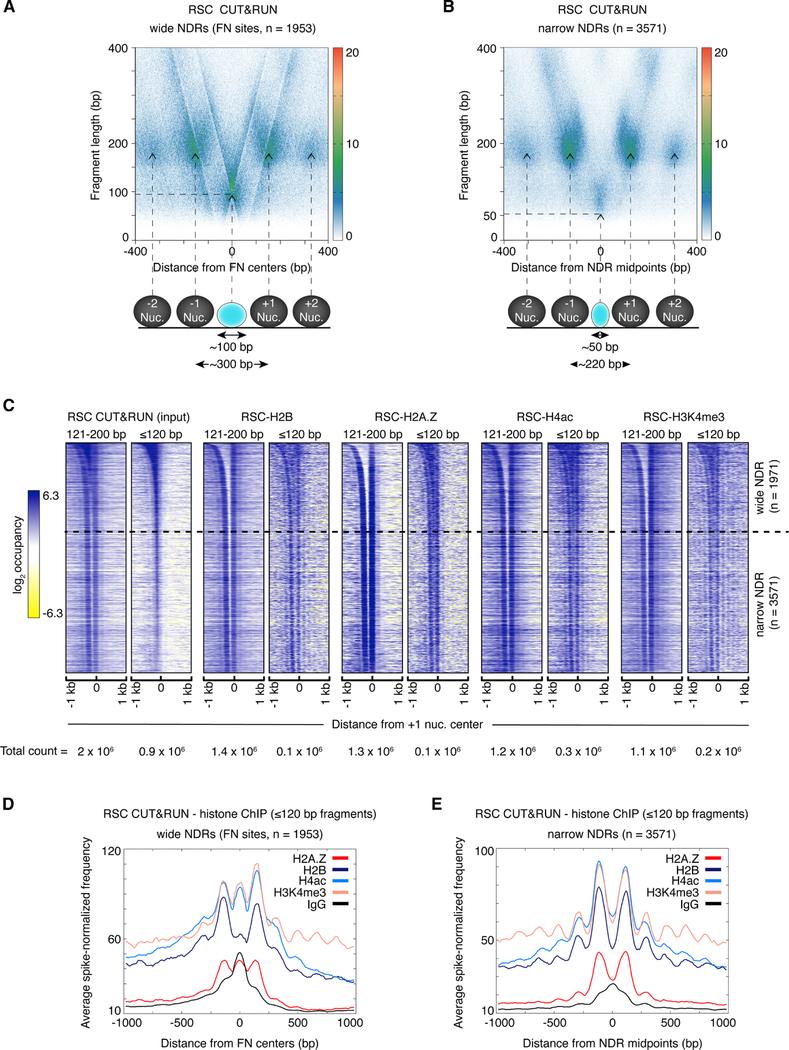
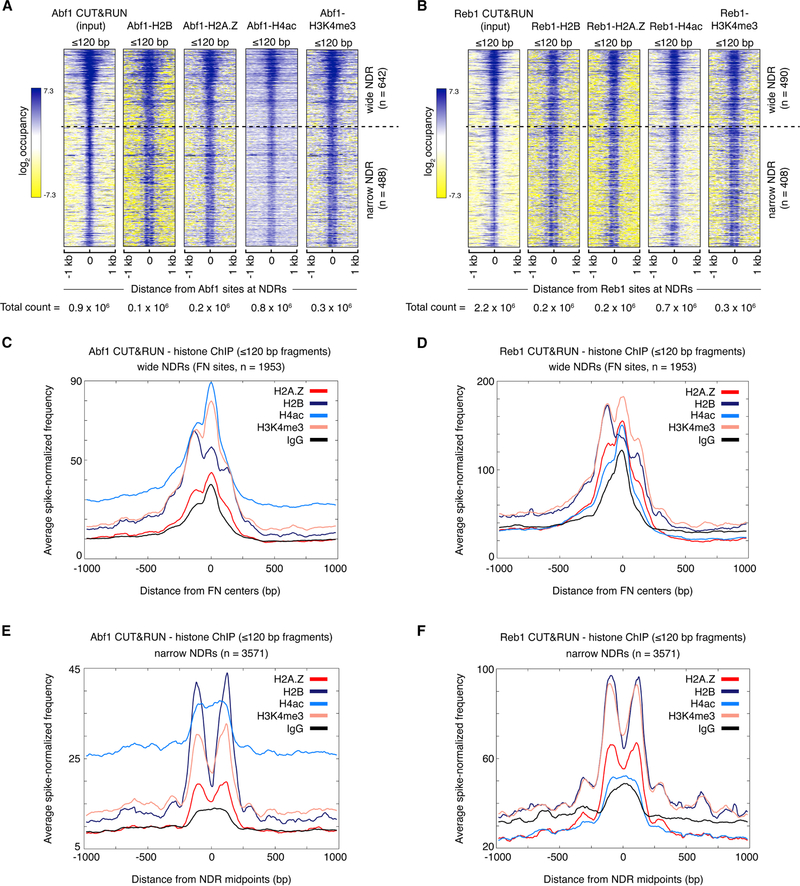
References
-
- Badis G, Chan ET, van Bakel H, Pena-Castillo L, Tillo D, Tsui K, Carlson CD, Gossett AJ, Hasinoff MJ, Warren CL, et al. (2008). A library of yeast transcription factor motifs reveals a widespread function for Rsc3 in targeting nucleosome exclusion at promoters. Molecular cell 32, 878–887. - PMC - PubMed
-
- Bernstein BE, Mikkelsen TS, Xie X, Kamal M, Huebert DJ, Cuff J, Fry B, Meissner A, Wernig M, Plath K, et al. (2006). A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell 125, 315–326. - PubMed
-
- Boeger H, Griesenbeck J, Strattan JS, and Kornberg RD (2003). Nucleosomes unfold completely at a transcriptionally active promoter. Molecular cell 11, 1587–1598. - PubMed
-
- Carrozza MJ, Li B, Florens L, Suganuma T, Swanson SK, Lee KK, Shia WJ, Anderson S, Yates J, Washburn MP, et al. (2005). Histone H3 methylation by Set2 directs deacetylation of coding regions by Rpd3S to suppress spurious intragenic transcription. Cell 123, 581–592. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
