

Key role of the REC lobe during CRISPR-Cas9 activation by 'sensing', 'regulating', and 'locking' the catalytic HNH domain
- PMID: 30555184
- PMCID: PMC6292676
- DOI: 10.1017/S0033583518000070
Key role of the REC lobe during CRISPR-Cas9 activation by 'sensing', 'regulating', and 'locking' the catalytic HNH domain
Abstract
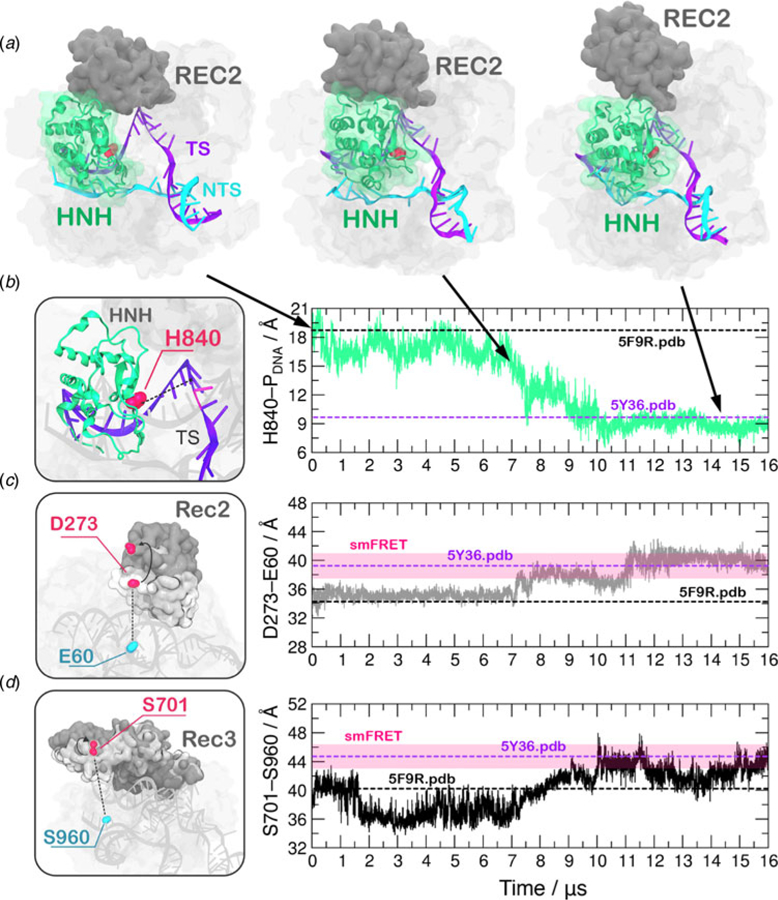
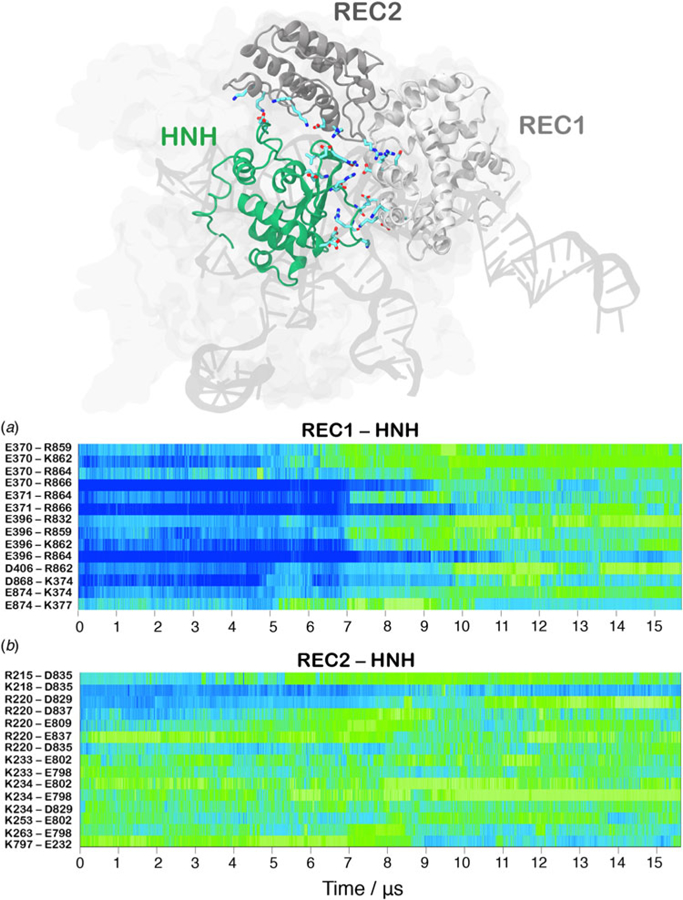
Understanding the conformational dynamics of CRISPR (clustered regularly interspaced short palindromic repeat)-Cas9 is of the utmost importance for improving its genome editing capability. Here, molecular dynamics simulations performed using Anton-2 - a specialized supercomputer capturing micro-to-millisecond biophysical events in real time and at atomic-level resolution - reveal the activation process of the endonuclease Cas9 toward DNA cleavage. Over the unbiased simulation, we observe that the spontaneous approach of the catalytic domain HNH to the DNA cleavage site is accompanied by a remarkable structural remodeling of the recognition (REC) lobe, which exerts a key role for DNA cleavage. Specifically, the significant conformational changes and the collective conformational dynamics of the REC lobe indicate a mechanism by which the REC1-3 regions 'sense' nucleic acids, 'regulate' the HNH conformational transition, and ultimately 'lock' the HNH domain at the cleavage site, contributing to its catalytic competence. By integrating additional independent simulations and existing experimental data, we provide a solid validation of the activated HNH conformation, which had been so far poorly characterized, and we deliver a comprehensive understanding of the role of REC1-3 in the activation process. Considering the importance of the REC lobe in the specificity of Cas9, this study poses the basis for fully understanding how the REC components control the cleavage of off-target sequences, laying the foundation for future engineering efforts toward improved genome editing.
Keywords: CRISPR–Cas9; genome editing; molecular dynamics; protein/nucleic acid interactions.
Conflict of interest statement
Conflict of interest. None.
Figures



References
-
- Abrams C and Bussi G (2014) Enhanced sampling in molecular dynamics using metadynamics, replica-exchange, and temperature-acceleration. Entropy 16, 163–199.
-
- Amadei A, Linssen ABM and Berendsen HJC (1993) Essential dynamics of proteins. Proteins-Structure Function and Genetics 17, 412–425. - PubMed
-
- Aqvist J (1990) Ion-water interaction potentials derived from free energy per-turbation simulations. Journal of Physical Chemistry 94, 8021–8024.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
