

Candida albicans β-Glucan Differentiates Human Monocytes Into a Specific Subset of Macrophages
- PMID: 30555483
- PMCID: PMC6284042
- DOI: 10.3389/fimmu.2018.02818
Candida albicans β-Glucan Differentiates Human Monocytes Into a Specific Subset of Macrophages
Abstract
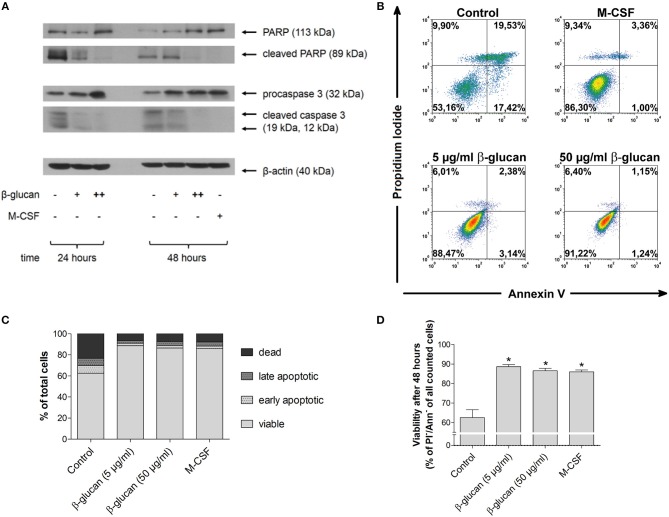
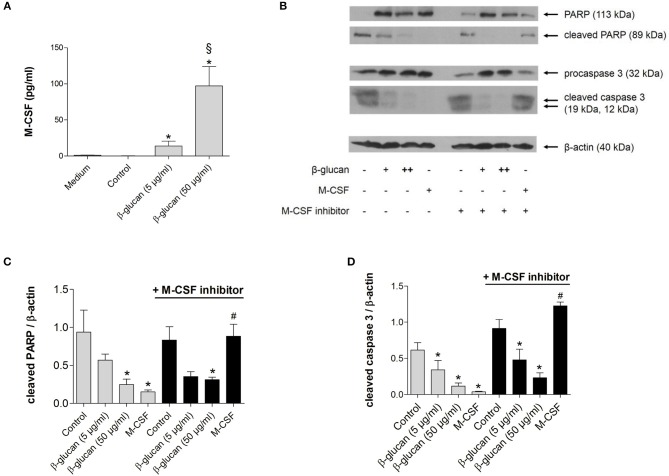
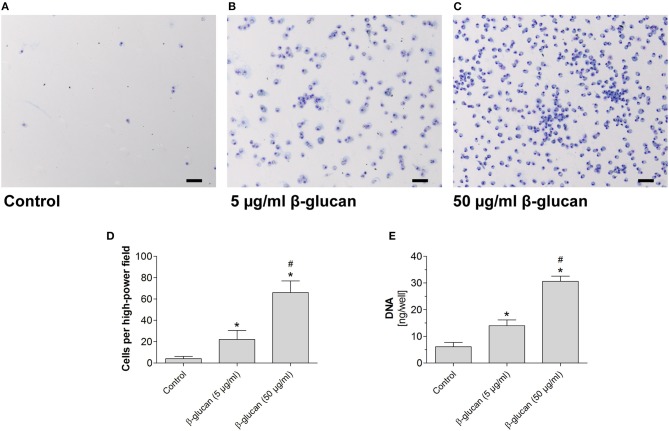
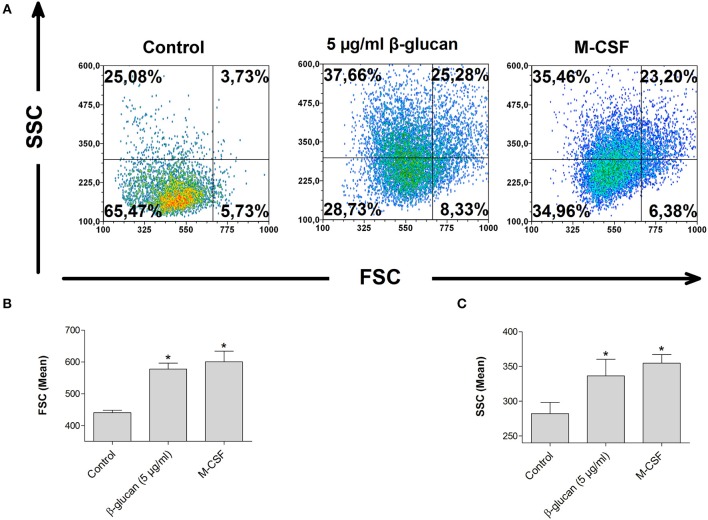
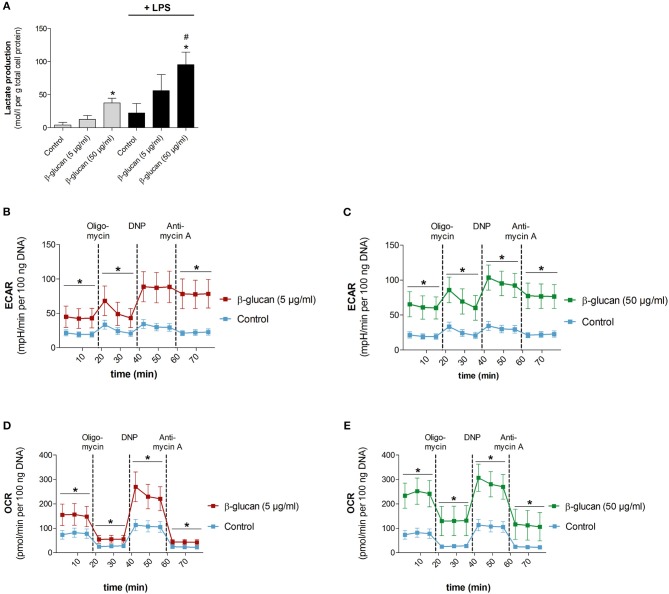
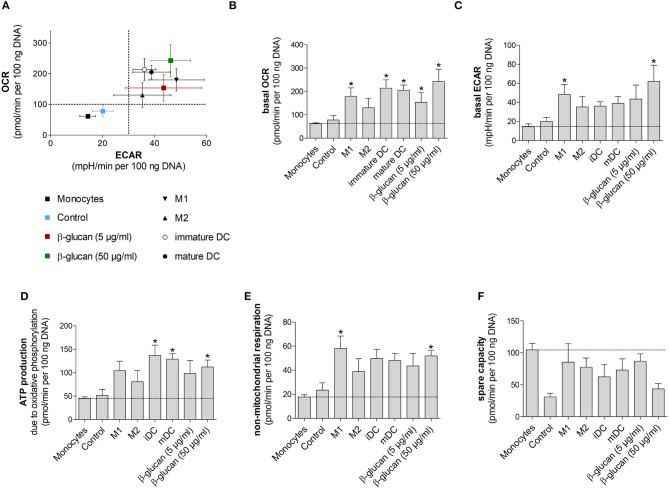
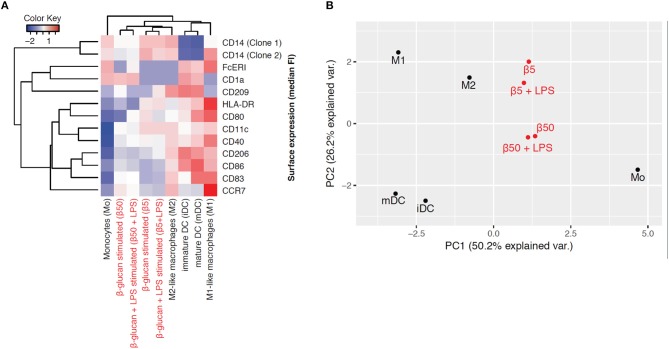
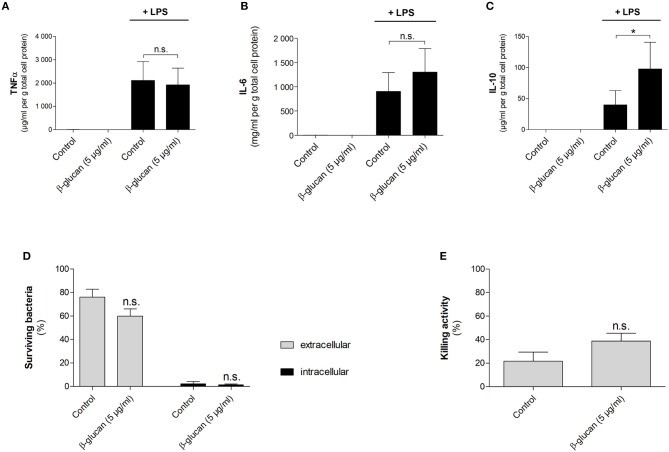
β-Glucan derived from cell walls of Candida albicans is a potent immune modulator. It has been shown to induce trained immunity in monocytes via epigenetic and metabolic reprogramming and to protect from lethal sepsis if applied prior to infection. Since β-glucan-trained monocytes have not been classified within the system of mononuclear phagocytes we analyzed these cells metabolically, phenotypically and functionally with a focus on monocyte-to-macrophage differentiation and compared them with naïve monocytes and other types of monocyte-derived cells such as classically (M1) or alternatively (M2) activated macrophages and monocyte-derived dendritic cells (moDCs). We show that β-glucan inhibits spontaneous apoptosis of monocytes independent from autocrine or paracrine M-CSF release and stimulates monocyte differentiation into macrophages. β-Glucan-differentiated macrophages exhibit increased cell size and granularity and enhanced metabolic activity when compared to naïve monocytes. Although β-glucan-primed cells expressed markers of alternative activation and secreted higher levels of IL-10 after lipopolysaccharide (LPS), their capability to release pro-inflammatory cytokines and to kill bacteria was unaffected. Our data demonstrate that β-glucan priming induces a population of immune competent long-lived monocyte-derived macrophages that may be involved in immunoregulatory processes.
Keywords: Candida albicans; monocyte survival; monocyte to macrophage differentiation; trained immunity; β-glucan.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials