

The Integrator complex regulates differential snRNA processing and fate of adult stem cells in the highly regenerative planarian Schmidtea mediterranea
- PMID: 30557303
- PMCID: PMC6312358
- DOI: 10.1371/journal.pgen.1007828
The Integrator complex regulates differential snRNA processing and fate of adult stem cells in the highly regenerative planarian Schmidtea mediterranea
Abstract
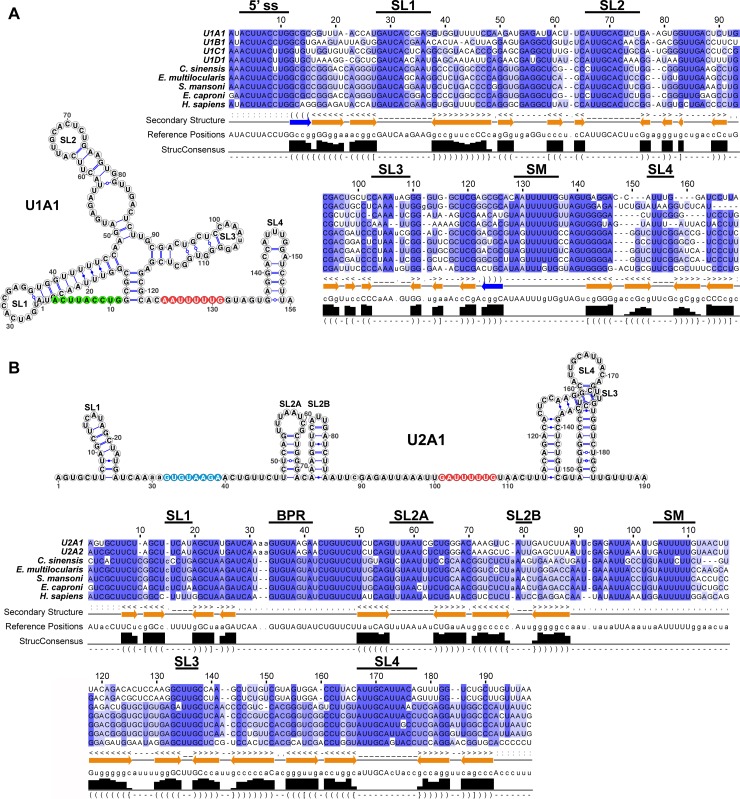
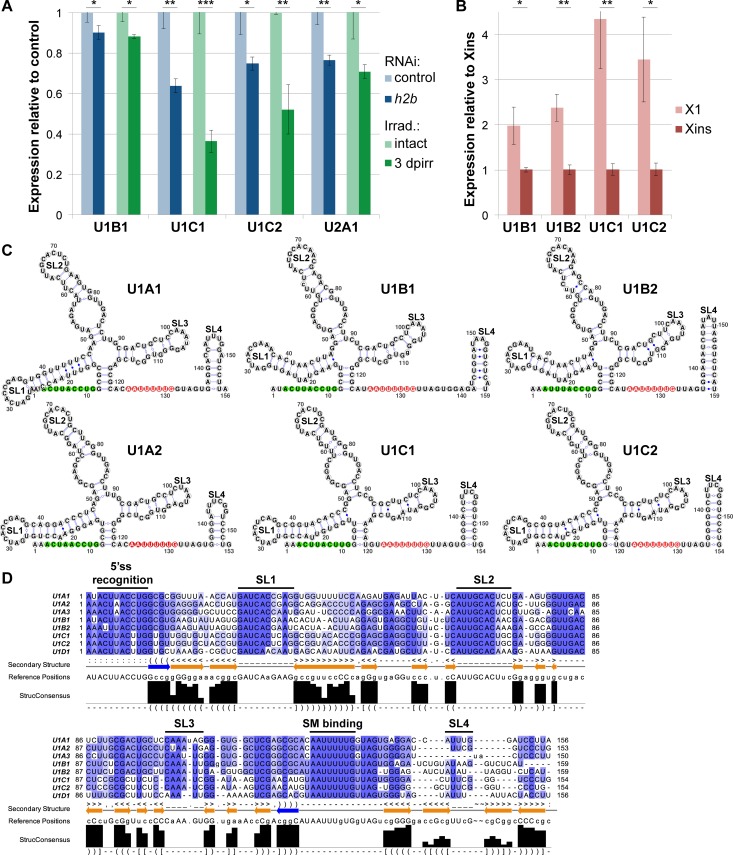
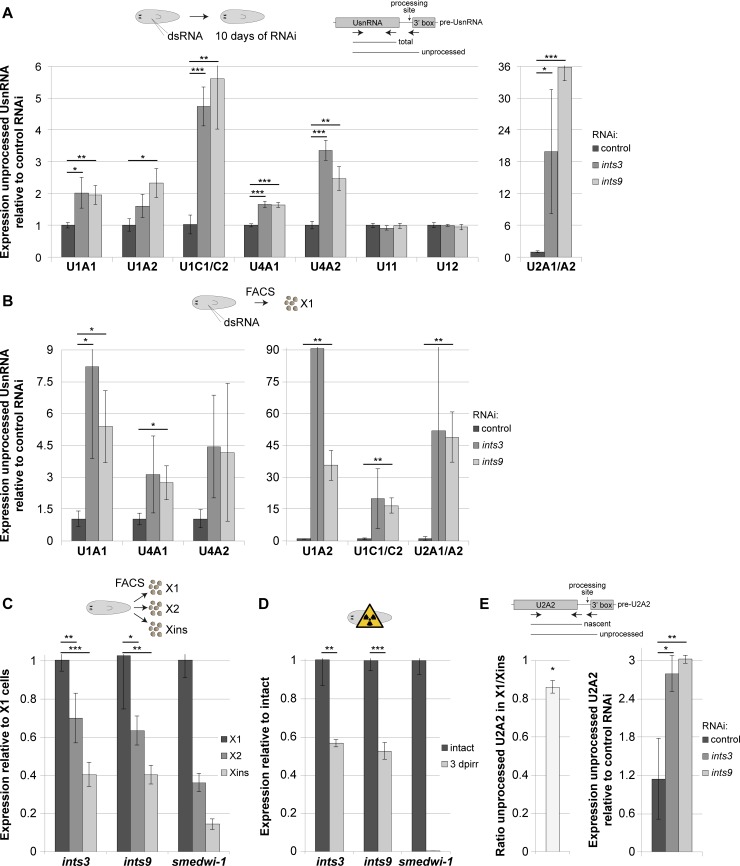
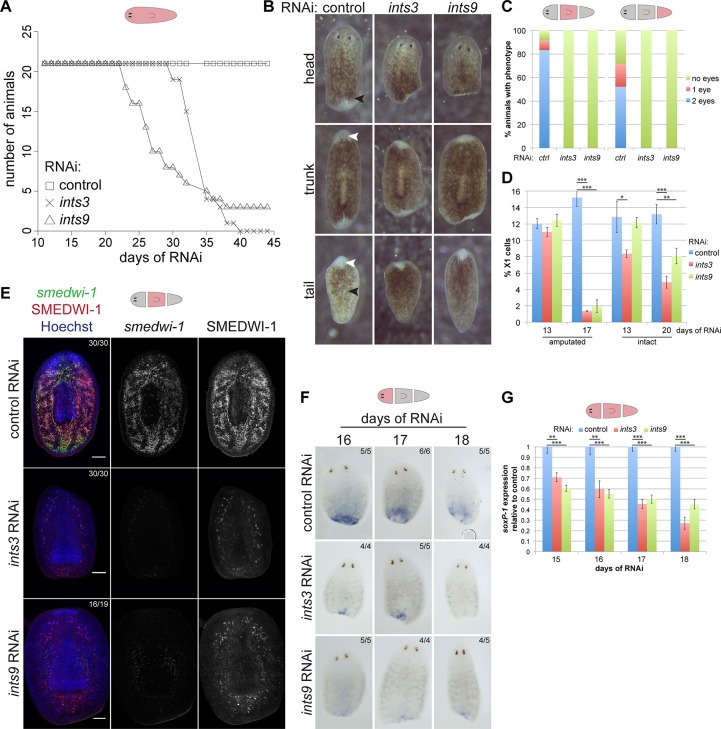
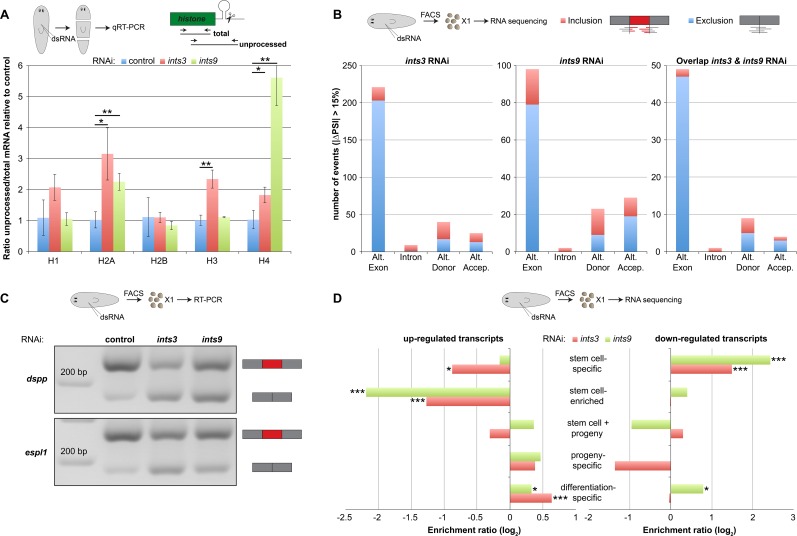
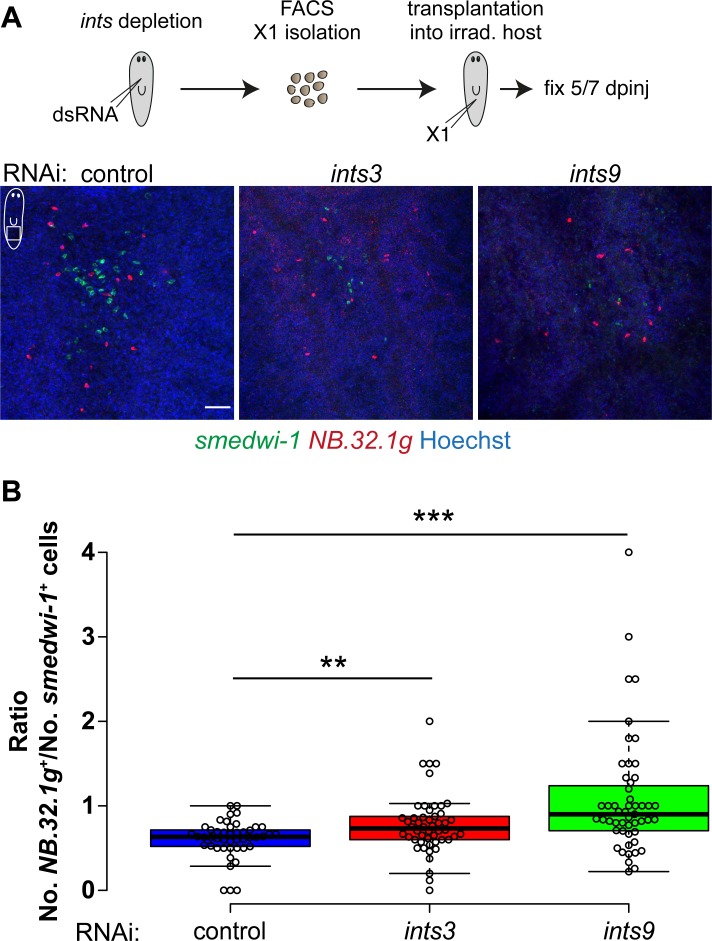
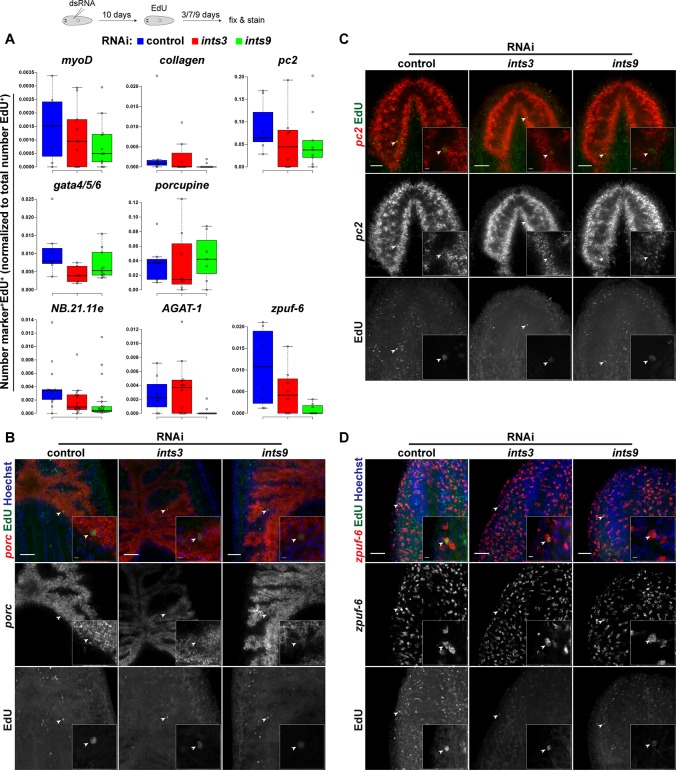
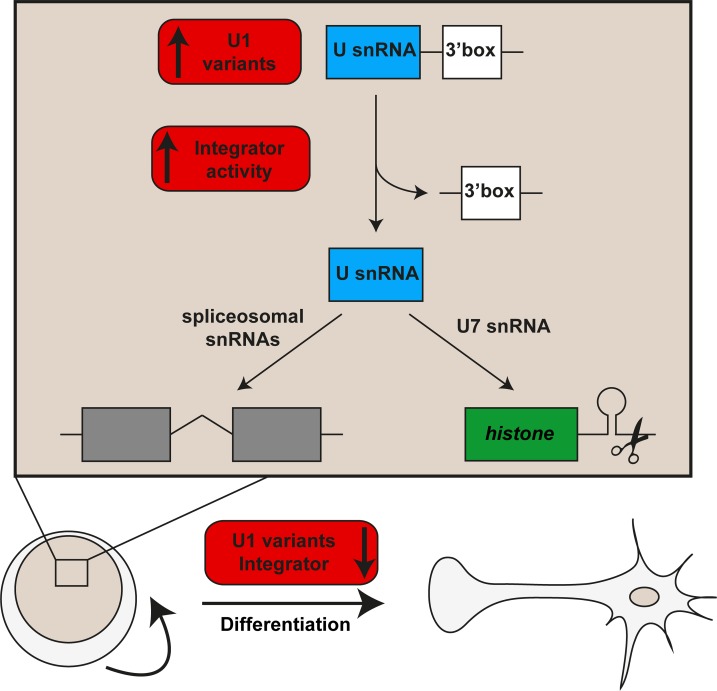
In multicellular organisms, cell type diversity and fate depend on specific sets of transcript isoforms generated by post-transcriptional RNA processing. Here, we used Schmidtea mediterranea, a flatworm with extraordinary regenerative abilities and a large pool of adult stem cells, as an in vivo model to study the role of Uridyl-rich small nuclear RNAs (UsnRNAs), which participate in multiple RNA processing reactions including splicing, in stem cell regulation. We characterized the planarian UsnRNA repertoire, identified stem cell-enriched variants and obtained strong evidence for an increased rate of UsnRNA 3'-processing in stem cells compared to their differentiated counterparts. Consistently, components of the Integrator complex showed stem cell-enriched expression and their depletion by RNAi disrupted UsnRNA processing resulting in global changes of splicing patterns and reduced processing of histone mRNAs. Interestingly, loss of Integrator complex function disrupted both stem cell maintenance and regeneration of tissues. Our data show that the function of the Integrator complex in UsnRNA 3'-processing is conserved in planarians and essential for maintaining their stem cell pool. We propose that cell type-specific modulation of UsnRNA composition and maturation contributes to in vivo cell fate choices, such as stem cell self-renewal in planarians.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Deep sequencing identifies new and regulated microRNAs in Schmidtea mediterranea.RNA. 2009 Aug;15(8):1483-91. doi: 10.1261/rna.1702009. Epub 2009 Jun 24. RNA. 2009. PMID: 19553344 Free PMC article.
-
Dissecting the function of Cullin-RING ubiquitin ligase complex genes in planarian regeneration.Dev Biol. 2018 Jan 15;433(2):210-217. doi: 10.1016/j.ydbio.2017.10.011. Epub 2017 Dec 25. Dev Biol. 2018. PMID: 29291974
-
Comprehensive annotation and characterization of planarian tRNA and tRNA-derived fragments (tRFs).RNA. 2021 Apr;27(4):477-495. doi: 10.1261/rna.077701.120. Epub 2021 Jan 14. RNA. 2021. PMID: 33446492 Free PMC article.
-
[Molecular approach to planarian stem cell system].Tanpakushitsu Kakusan Koso. 2005 May;50(6 Suppl):706-10. Tanpakushitsu Kakusan Koso. 2005. PMID: 15926503 Review. Japanese. No abstract available.
-
Planarian regeneration: its end is its beginning.Cell. 2006 Jan 27;124(2):241-5. doi: 10.1016/j.cell.2006.01.012. Cell. 2006. PMID: 16439195 Review.
Cited by
-
Efficient depletion of ribosomal RNA for RNA sequencing in planarians.BMC Genomics. 2019 Nov 29;20(1):909. doi: 10.1186/s12864-019-6292-y. BMC Genomics. 2019. PMID: 31783730 Free PMC article.
-
Genomic regulation of transcription and RNA processing by the multitasking Integrator complex.Nat Rev Mol Cell Biol. 2023 Mar;24(3):204-220. doi: 10.1038/s41580-022-00534-2. Epub 2022 Sep 30. Nat Rev Mol Cell Biol. 2023. PMID: 36180603 Free PMC article. Review.
-
The Integrator complex at the crossroad of coding and noncoding RNA.Curr Opin Cell Biol. 2021 Jun;70:37-43. doi: 10.1016/j.ceb.2020.11.003. Epub 2020 Dec 16. Curr Opin Cell Biol. 2021. PMID: 33340967 Free PMC article. Review.
-
The exon junction complex is required for stem and progenitor cell maintenance in planarians.Dev Biol. 2020 Jan 1;457(1):119-127. doi: 10.1016/j.ydbio.2019.09.010. Epub 2019 Sep 23. Dev Biol. 2020. PMID: 31557470 Free PMC article.
-
INTS11 regulates hematopoiesis by promoting PRC2 function.Sci Adv. 2021 Sep 3;7(36):eabh1684. doi: 10.1126/sciadv.abh1684. Epub 2021 Sep 1. Sci Adv. 2021. PMID: 34516911 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
