

Diverse members of the Xylariales lack canonical mating-type regions
- PMID: 30557613
- PMCID: PMC6321786
- DOI: 10.1016/j.fgb.2018.12.004
Diverse members of the Xylariales lack canonical mating-type regions
Abstract
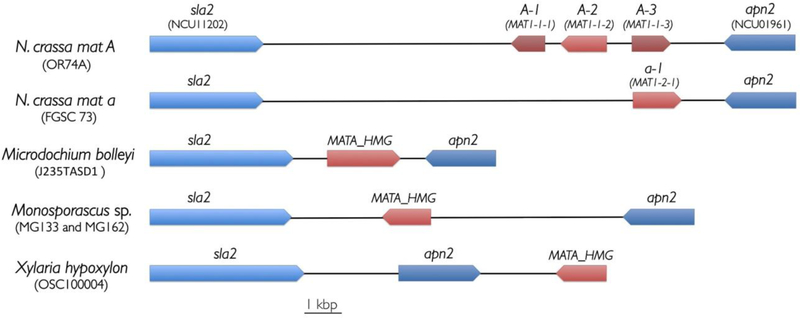
A survey of genomes reported here for 10 isolates of Monosporascus species and an additional 25 genomes from other members of the Xylariales (representing 15 genera) available in public databases indicated that genes typically associated with MAT1-1 (mat A) or MAT1-2 (mat a) mating types are absent or have diverged greatly relative to counterparts in other Pezizomycotina. This was particularly surprising for isolates known to be homothallic, given that homothallic members of the Pezizomycotina typically possess a MAT1-1-1 (mat A-1) gene and one or both of two other closely-linked mating-type genes, MAT1-1-2 (mat A-2) and MAT1-1-3 (mat A-3), in addition to MAT1-2-1 (mat a-1). We failed to detect candidate genes for either MAT1-1-1 or MAT1-1-2 in any member of the Xylariales. Genes related to MAT1-2-1 and MAT1-1-3 are present in the genomes examined, but most appear to be orthologs of MATA_HMG (high-mobility group) genes with non-mating-type functions rather than orthologs of mating-type genes. Several MATA_HMG genes were found in genome positions that suggest they are derived from mating-type genes, but these genes are highly divergent relative to known MAT1-2-1 and MAT1-1-3 genes. The genomes examined represent substantial diversity within the order and include M. cannonballus, M. ibericus, Xylaria hypoxylon, X. striata, Daldinia eschscholzii, Eutypa lata, Rosellinia necatrix, Microdochium bolleyi and several others. We employed a number of avenues to search for homologs, including multiple BLAST approaches and examination of annotated genes adjacent to genes known to flank mating regions in other members of the Ascomycota. The results suggest that the mating regions have been lost from, or altered dramatically in, the Xylariales genomes examined and that mating and sexual development in these fungi are controlled differently than has been reported for members of the Pezizomycotina studied to date.
Keywords: Genomics; Mating type; Microdochium; Monosporascus; Xylaria; Xylariales.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Characterization and phylogenetic analysis of the mating-type loci in the asexual ascomycete genus Ulocladium.Mycologia. 2014 Jul-Aug;106(4):649-65. doi: 10.3852/13-383. Epub 2014 Jun 2. Mycologia. 2014. PMID: 24891417
-
Cloning and characterization of the mating type (MAT) locus from Ascochyta rabiei (teleomorph: Didymella rabiei) and a MAT phylogeny of legume-associated Ascochyta spp.Fungal Genet Biol. 2003 Jul;39(2):151-67. doi: 10.1016/s1087-1845(03)00015-x. Fungal Genet Biol. 2003. PMID: 12781674
-
The novel Huntiella omanensis mating gene, MAT1-2-7, is essential for ascomatal maturation.Fungal Genet Biol. 2020 Apr;137:103335. doi: 10.1016/j.fgb.2020.103335. Epub 2020 Jan 17. Fungal Genet Biol. 2020. PMID: 31958567
-
[Classification of dermatophytes by mating type (MAT) gene analysis].Med Mycol J. 2012;53(3):175-8. doi: 10.3314/mmj.53.175. Med Mycol J. 2012. PMID: 23149351 Review. Japanese.
-
Genetic Networks That Govern Sexual Reproduction in the Pezizomycotina.Microbiol Mol Biol Rev. 2021 Dec 15;85(4):e0002021. doi: 10.1128/MMBR.00020-21. Epub 2021 Sep 29. Microbiol Mol Biol Rev. 2021. PMID: 34585983 Free PMC article. Review.
Cited by
-
Importance of appropriate genome information for the design of mating type primers in black and yellow morel populations.IMA Fungus. 2022 Aug 22;13(1):14. doi: 10.1186/s43008-022-00101-6. IMA Fungus. 2022. PMID: 35996182 Free PMC article.
-
Genomic Analysis of Diverse Members of the Fungal Genus Monosporascus Reveals Novel Lineages, Unique Genome Content and a Potential Bacterial Associate.G3 (Bethesda). 2020 Aug 5;10(8):2573-2583. doi: 10.1534/g3.120.401489. G3 (Bethesda). 2020. PMID: 32580939 Free PMC article.
-
Population genomics of the grapevine pathogen Eutypa lata reveals evidence for population expansion and intraspecific differences in secondary metabolite gene clusters.PLoS Genet. 2022 Apr 1;18(4):e1010153. doi: 10.1371/journal.pgen.1010153. eCollection 2022 Apr. PLoS Genet. 2022. PMID: 35363788 Free PMC article.
References
-
- Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, Kulikov AS, Lesin VM, Nikolenko SI, Pham S, Prjibelski AD, Pyshkin AV, Sirotkin AV, Vyahhi N, Tesler G, Alekseyev MA, Pevzner PA, 2012. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol 19, 455–477. - PMC - PubMed
-
- Ben Salem I, Correia KC, Boughalleb N, Michereff SJ, Leon M, Abad-Campos P, Garcia-Jimenez J, Armengol J, 2013. Monosporascus eutypoides, a cause of root rot and vine decline in Tunisia, and evidence that M. cannonballus and M. eutypoides are distinct species. Plant Disease 97, 737–743 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
