

Retinal degeneration 3 (RD3) protein, a retinal guanylyl cyclase regulator, forms a monomeric and elongated four-helix bundle
- PMID: 30559291
- PMCID: PMC6378972
- DOI: 10.1074/jbc.RA118.006106
Retinal degeneration 3 (RD3) protein, a retinal guanylyl cyclase regulator, forms a monomeric and elongated four-helix bundle
Abstract
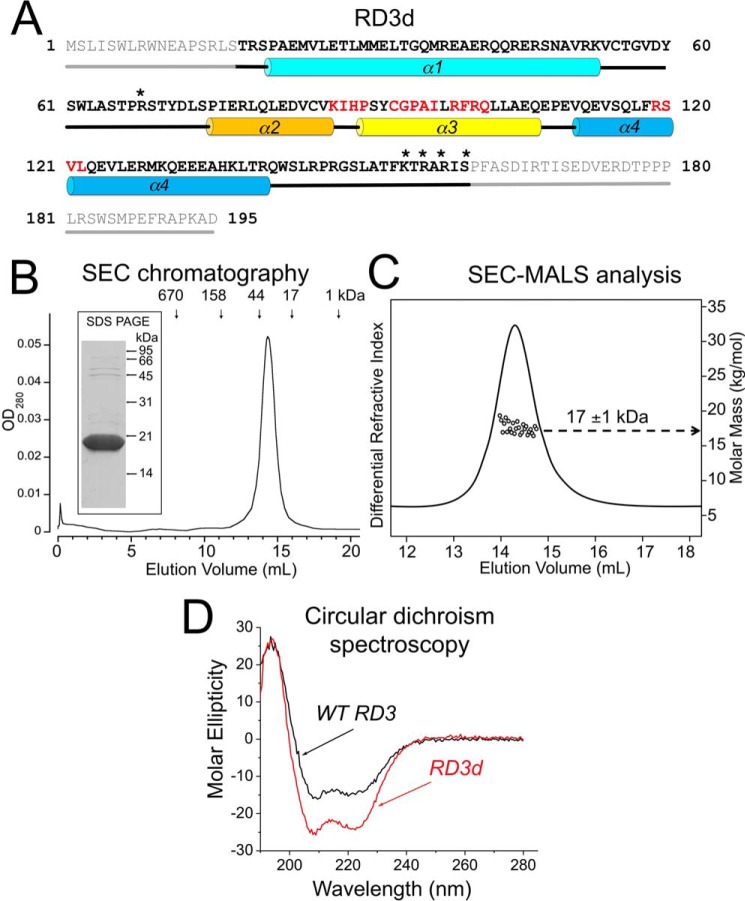
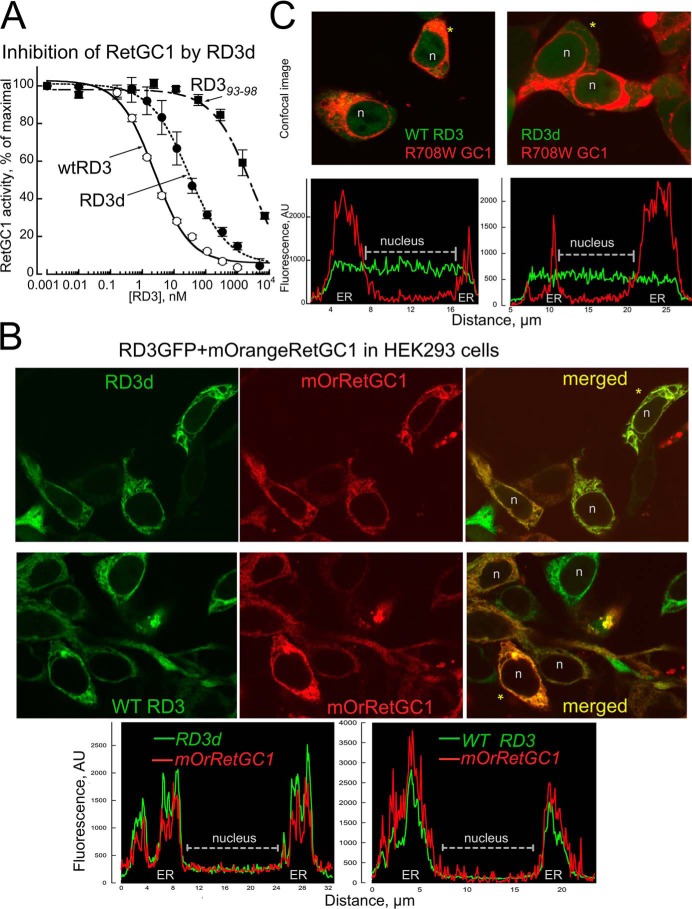
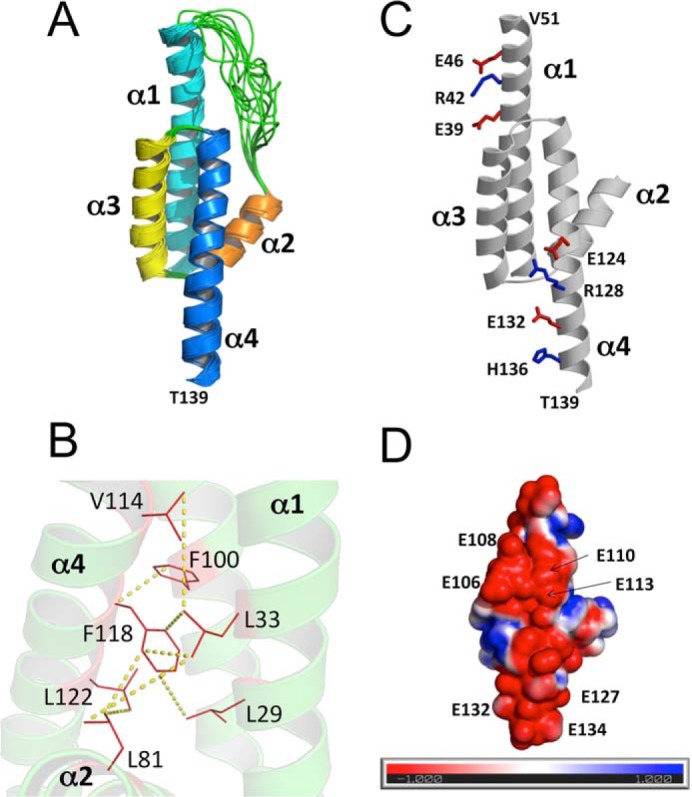
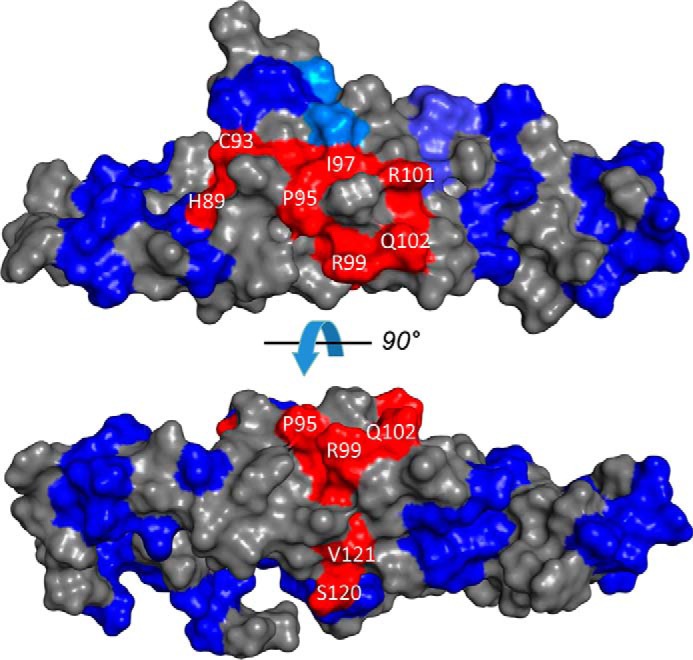
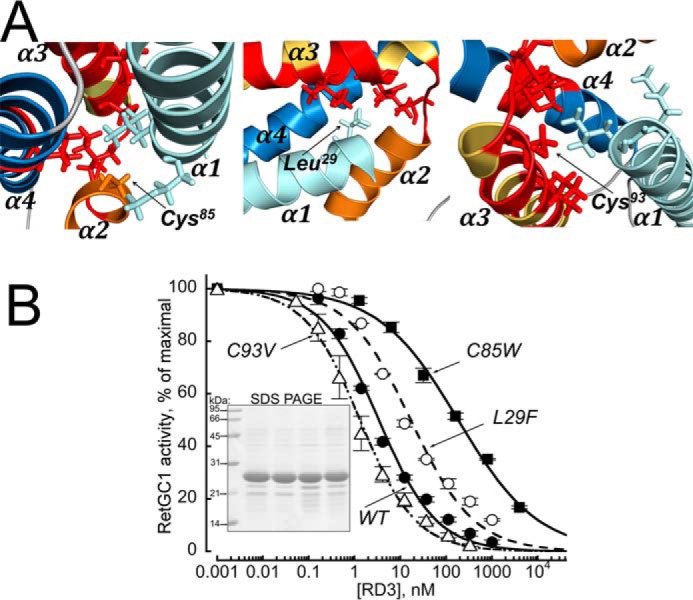
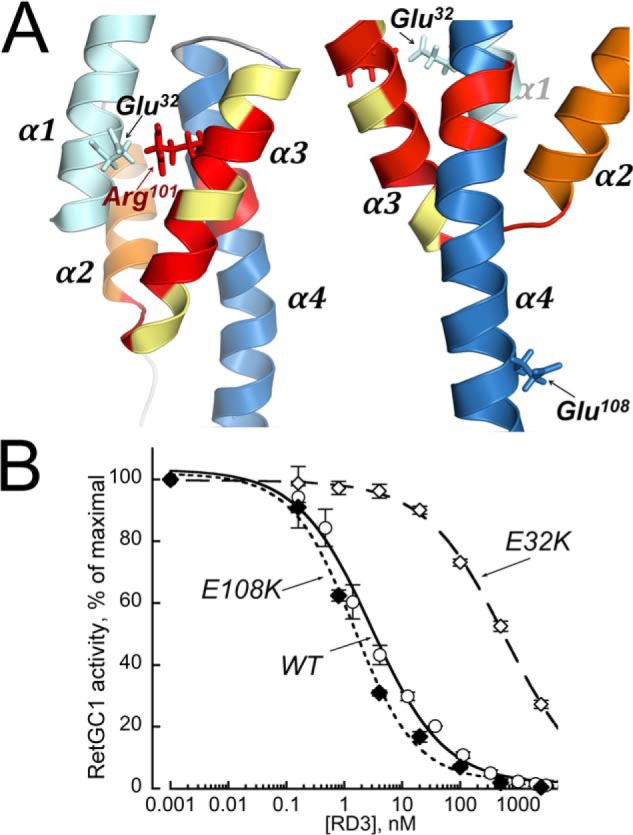
Retinal degeneration 3 (RD3) protein promotes accumulation of retinal membrane guanylyl cyclase (RetGC) in the photoreceptor outer segment and suppresses RetGC activation by guanylyl cyclase-activating proteins (GCAPs). Mutations truncating RD3 cause severe congenital blindness by preventing the inhibitory binding of RD3 to the cyclase. The high propensity of RD3 to aggregate in solution has prevented structural analysis. Here, we produced a highly soluble variant of human RD3 (residues 18-160) that is monomeric and can still bind and negatively regulate RetGC. The NMR solution structure of RD3 revealed an elongated backbone structure (70 Å long and 30 Å wide) consisting of a four-helix bundle with a long unstructured loop between helices 1 and 2. The structure reveals that RD3 residues previously implicated in the RetGC binding map to a localized and contiguous area on the structure, involving a loop between helices 2 and 3 and adjacent parts of helices 3 and 4. The NMR structure of RD3 was validated by mutagenesis. Introducing Trp85 or Phe29 to replace Cys or Leu, respectively, disrupts packing in the hydrophobic core and lowers RD3's apparent affinity for RetGC1. Introducing a positive charge at the interface (Glu32 to Lys) also lowered the affinity. Conversely, introducing Val in place of Cys93 stabilized the hydrophobic core and increased the RD3 affinity for the cyclase. The NMR structure of RD3 presented here provides a structural basis for elucidating RD3-RetGC interactions relevant for normal vision or blindness.
Keywords: guanylate cyclase (guanylyl cyclase); nuclear magnetic resonance (NMR); photoreceptor; phototransduction; retinal degeneration.
© 2019 Peshenko et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
