

Novel Widespread Marine Oomycetes Parasitising Diatoms, Including the Toxic Genus Pseudo-nitzschia: Genetic, Morphological, and Ecological Characterisation
- PMID: 30559730
- PMCID: PMC6286980
- DOI: 10.3389/fmicb.2018.02918
Novel Widespread Marine Oomycetes Parasitising Diatoms, Including the Toxic Genus Pseudo-nitzschia: Genetic, Morphological, and Ecological Characterisation
Abstract
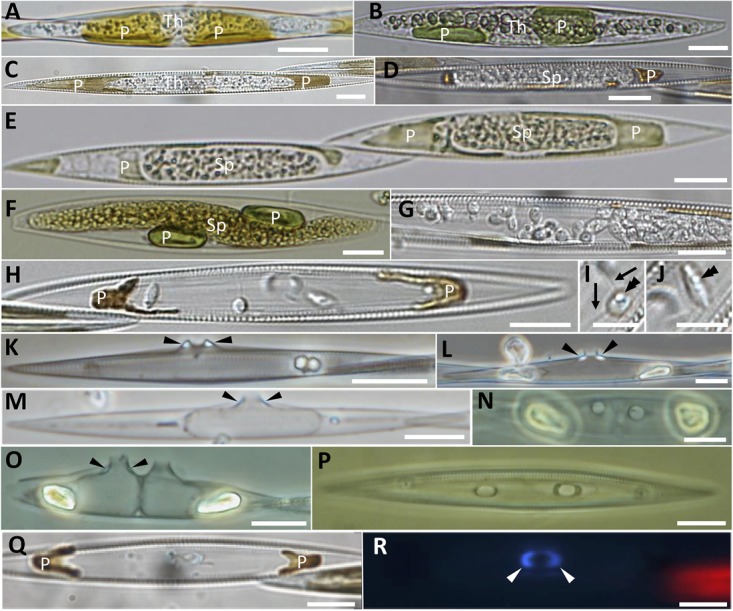
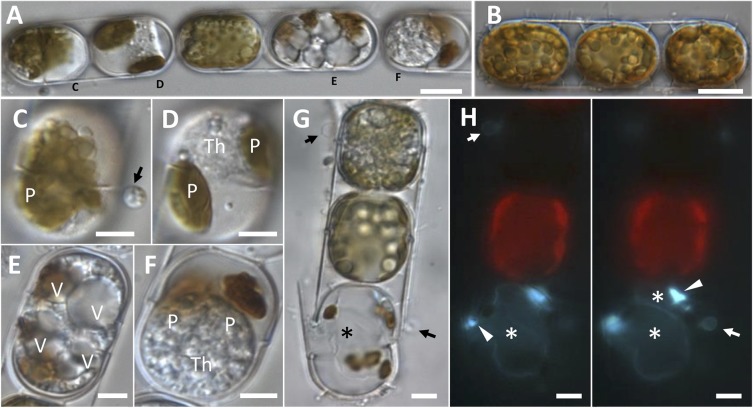
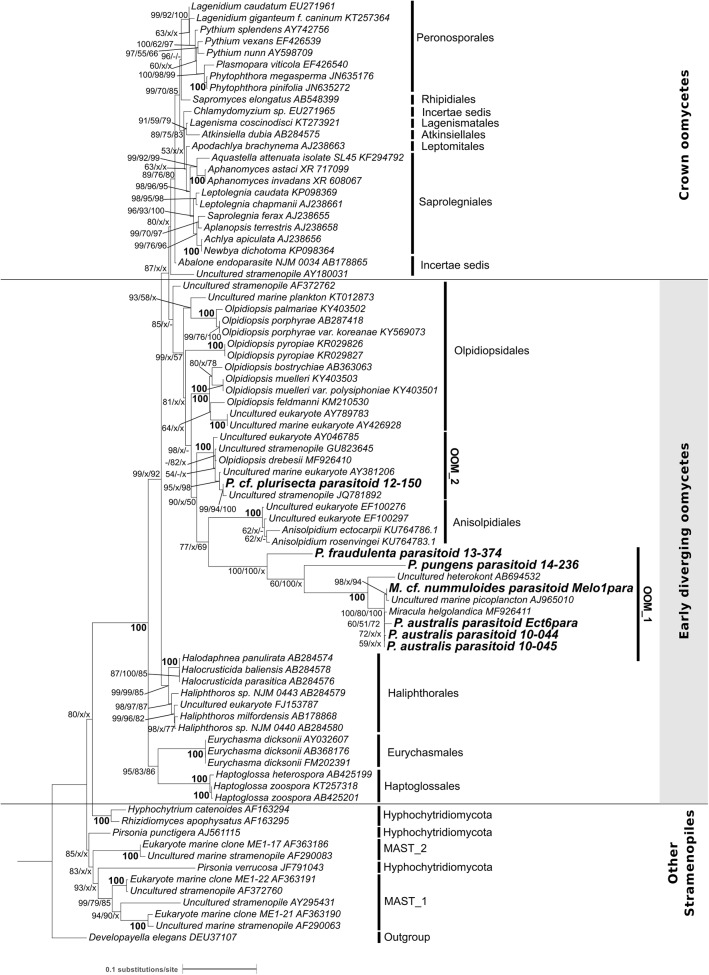
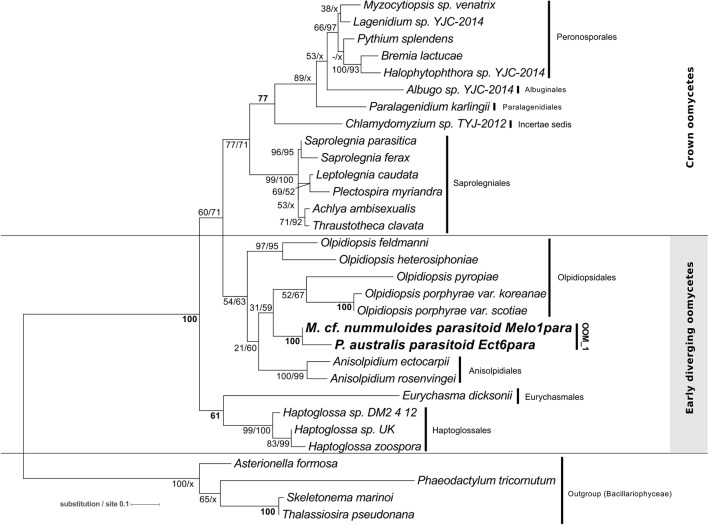
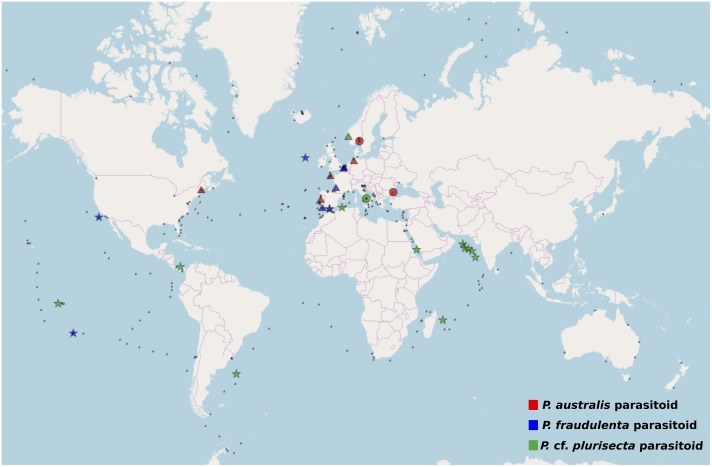
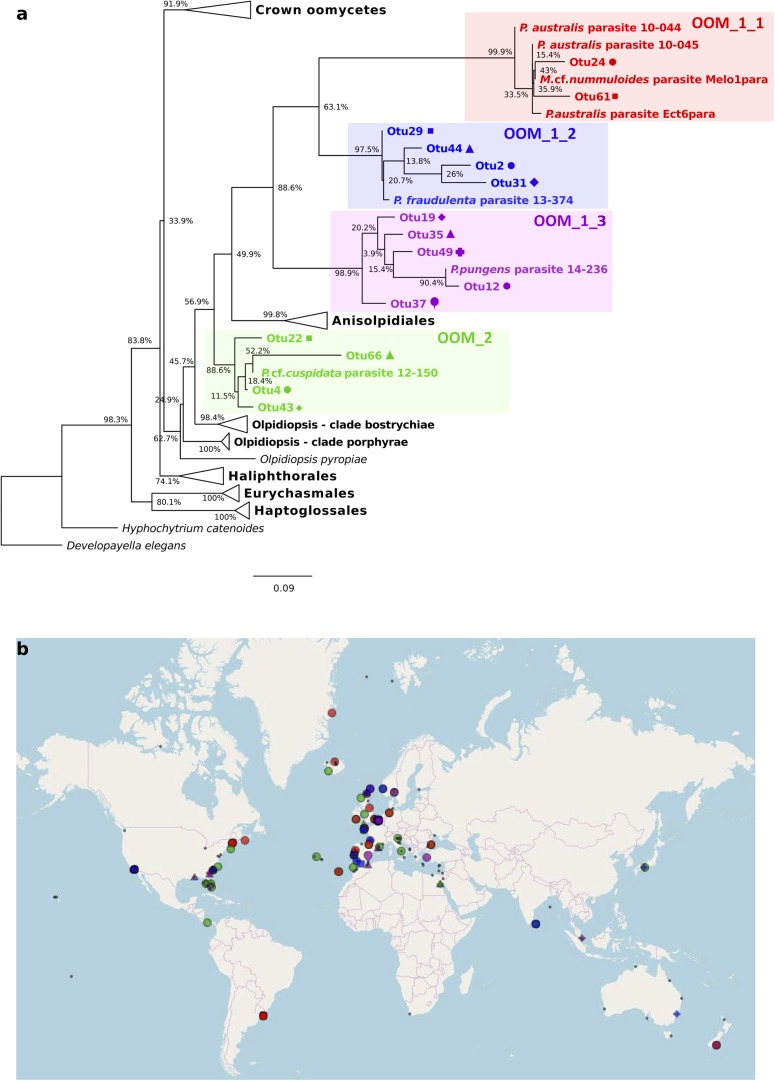
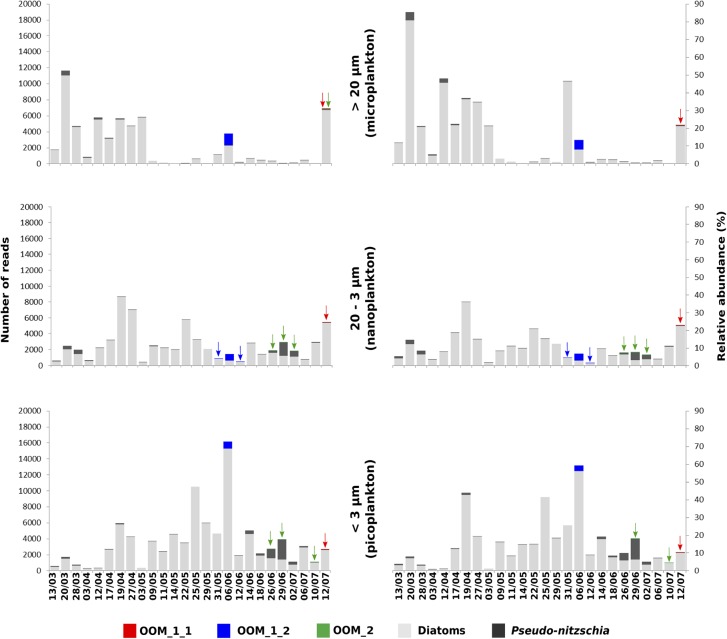
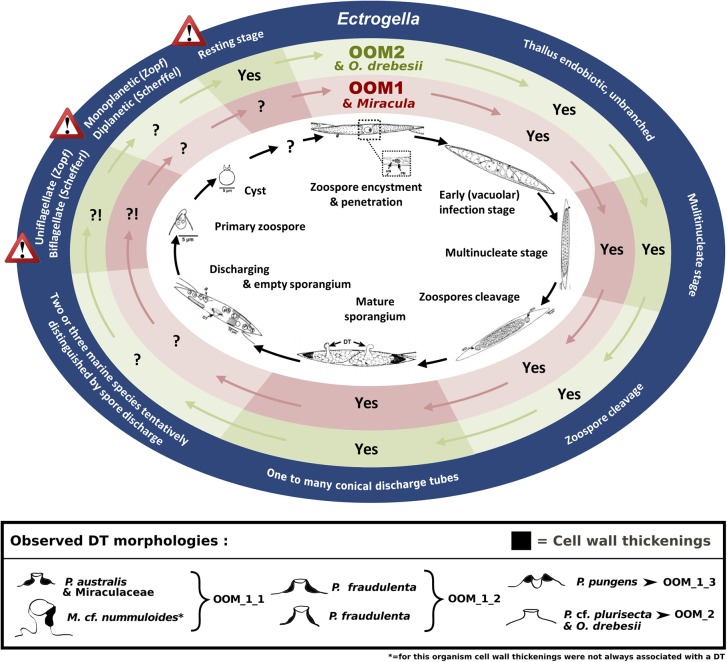
Parasites are key drivers of phytoplankton bloom dynamics and related aquatic ecosystem processes. Yet, the dearth of morphological and molecular information hinders the assessment of their diversity and ecological role. Using single-cell techniques, we characterise morphologically and molecularly, intracellular parasitoids infecting four potentially toxin-producing Pseudo-nitzschia and one Melosira species on the North Atlantic coast. These sequences define two, morphologically indistinguishable clades within the phylum Oomycota, related to the genera of algal parasites Anisolpidium and Olpidiopsis and the diatom parasitoid species Miracula helgolandica. Our morphological data are insufficient to attribute either clade to the still unsequenced genus Ectrogella; hence it is proposed to name the clades OOM_1 and OOM_2. A screening of global databases of the barcode regions V4 and V9 of the 18S rDNA demonstrate the presence of these parasitoids beyond the North Atlantic coastal region. During a biweekly metabarcoding survey (Concarneau Bay, France), reads associated with one sequenced parasitoid coincided with the decline of Cerataulina pelagica bloom, whilst the other parasitoids co-occurred at low abundance with Pseudo-nitzschia. Our data highlight a complex and unexplored diversity of the oomycete parasitoids of diatoms and calls for the investigation of their phenology, evolution, and potential contribution in controlling their host spatial-temporal dynamics.
Keywords: Pseudo-nitzschia; diatoms; marine oomycetes; metabarcoding; plankton parasites; single-cell analysis.
Figures
References
-
- Alster A., Zohary T. (2007). Interactions between the bloom-forming dinoflagellate Peridinium gatunense and the chytrid fungus Phlyctochytrium sp. Hydrobiologia 578 131–139. 10.1007/s10750-006-0439-y - DOI
-
- Beakes G. W., Honda D., Thines M. (2014). “Systematics of the straminipila: labyrinthulomycota, hyphochytriomycota, and oomycota,” in Systematics and Evolution, 2nd Edn, eds McLaughlin D. J., Spatafora J. W. (Berlin: Springer-Verlag; ), 39–97.
-
- Beakes G. W., Thines M. (2016). “Hyphochytriomycota and oomycota,” in Handbook of the Protists, eds Archibald J. M., Simpson A. G. B., Slamovits C. (Cham: Springer International Publishing; ), 3–71.
LinkOut - more resources
Full Text Sources