

The occurrence and formation of monoterpenes in herbivore-damaged poplar roots
- PMID: 30560919
- PMCID: PMC6299004
- DOI: 10.1038/s41598-018-36302-6
The occurrence and formation of monoterpenes in herbivore-damaged poplar roots
Abstract
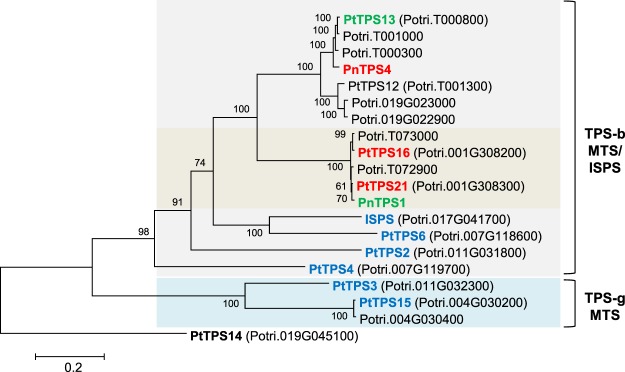
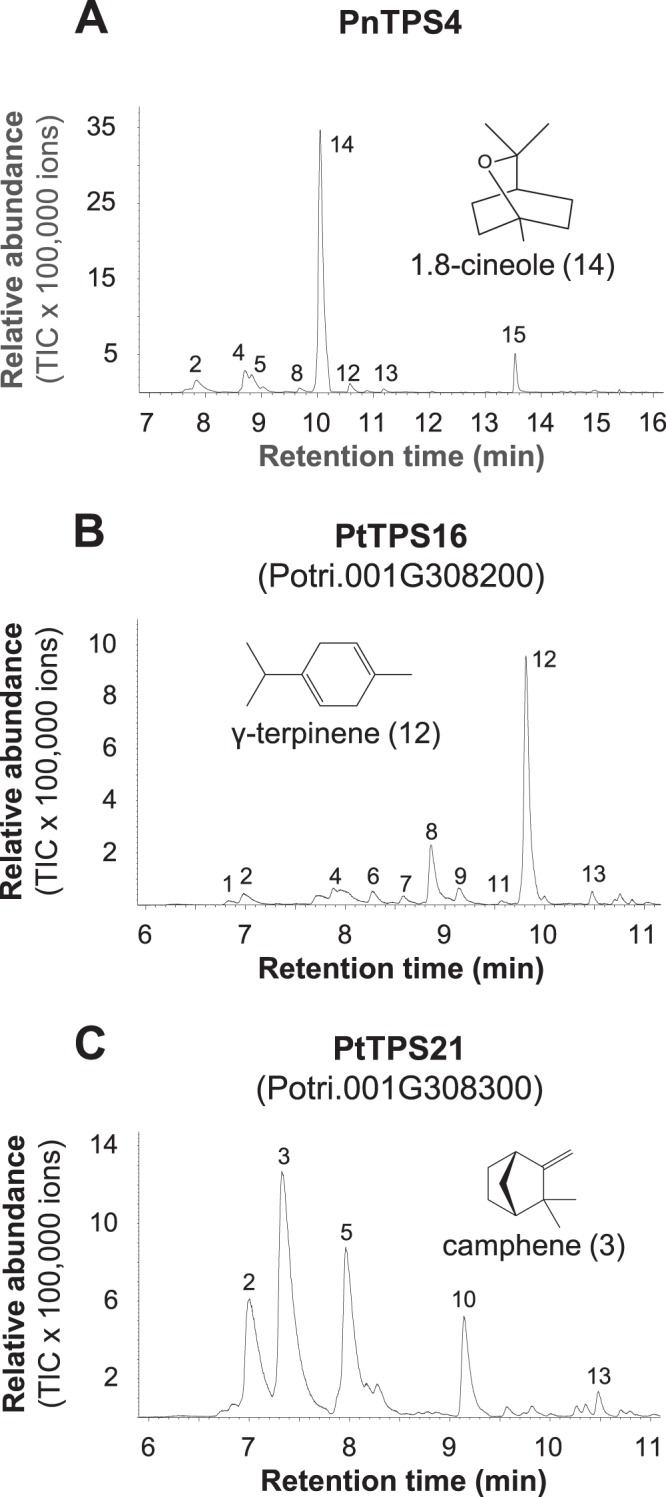
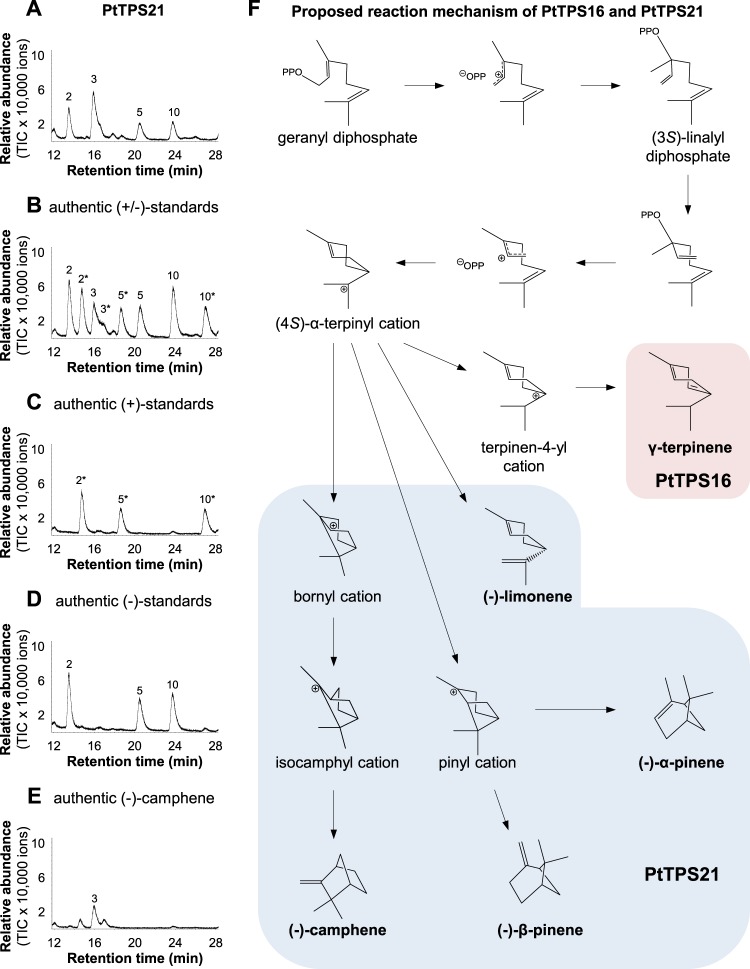
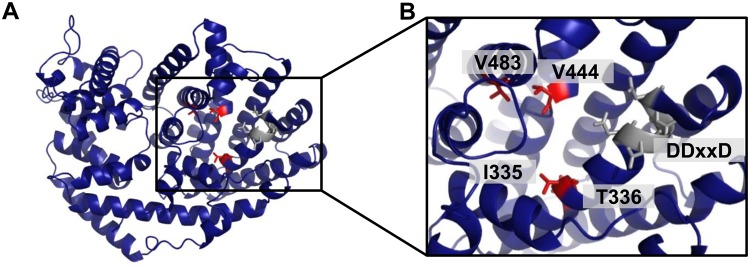
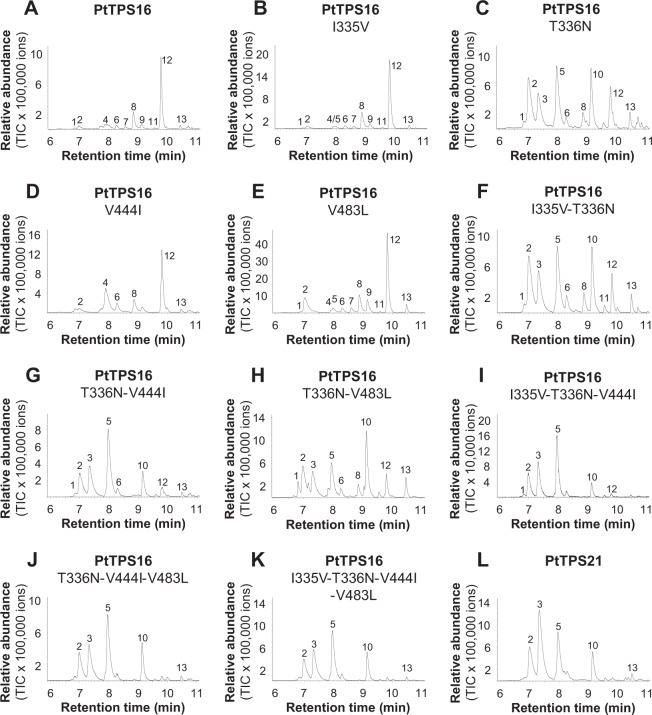
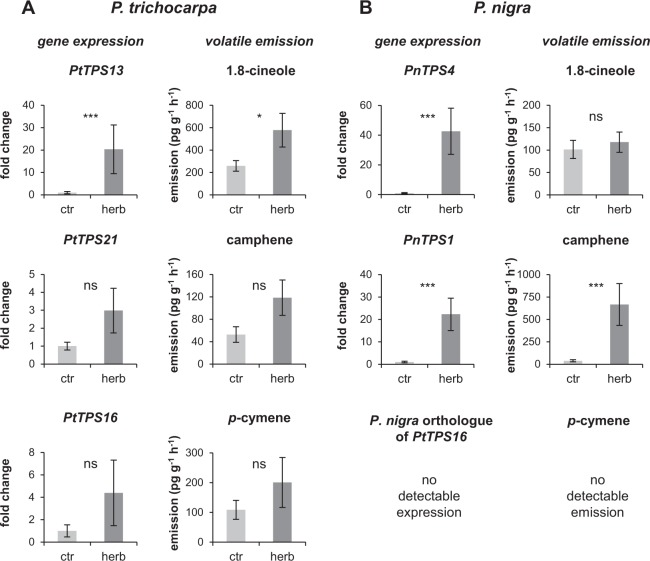
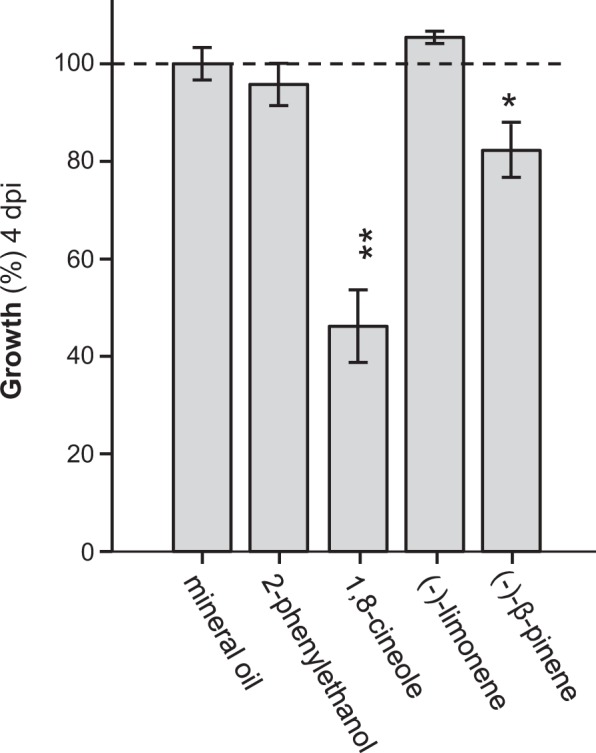
Volatiles are often released upon herbivory as plant defense compounds. While the formation of volatiles above-ground has been intensively studied, little is known about herbivore-induced root volatiles. Here, we show that cockchafer larvae-damaged roots of Populus trichocarpa and P. nigra release a mixture of monoterpenes, including (-)-α-pinene, (-)-camphene, (-)-β-pinene, p-cymene, and 1,8-cineole. Three terpene synthases, PtTPS16 and PtTPS21 from P. trichocarpa and PnTPS4 from P. nigra, could be identified and characterized in vitro. PnTPS4 was found to produce 1,8-cineole as sole product. PtTPS16 and PtTPS21, although highly similar to each other, showed different product specificities and produced γ-terpinene and a mixture of (-)-camphene, (-)-α-pinene, (-)-β-pinene, and (-)-limonene, respectively. Four active site residues were found to determine the different product specificities of the two enzymes. The expression profiles of PtTPS16, PtTPS21, and PnTPS4 in undamaged and herbivore-damaged poplar roots generally matched the emission pattern of monoterpenes, indicating that monoterpene emission in roots is mainly determined at the gene transcript level. Bioassays with Phytophtora cactorum (Oomycetes) revealed inhibitory effects of vapor-phase 1,8-cineole and (-)-β-pinene on the growth of this important plant pathogen. Thus herbivore-induced volatile monoterpenes may have a role in defense against pathogens that cause secondary infections after root wounding.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Quintana-Rodriguez E, Rivera-Macias LE, Adame-Alvarez RM, Torres JM, Heil M. Shared weapons in fungus-fungus and fungus-plant interactions? Volatile organic compounds of plant or fungal origin exert direct antifungal activity in vitro. Fungal Ecology. 2018;33:115–121. doi: 10.1016/j.funeco.2018.02.005. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
