

Arsenic modifies serotonin metabolism through glucuronidation in pancreatic β-cells
- PMID: 30562058
- PMCID: PMC6459295
- DOI: 10.1152/ajpendo.00302.2018
Arsenic modifies serotonin metabolism through glucuronidation in pancreatic β-cells
Abstract
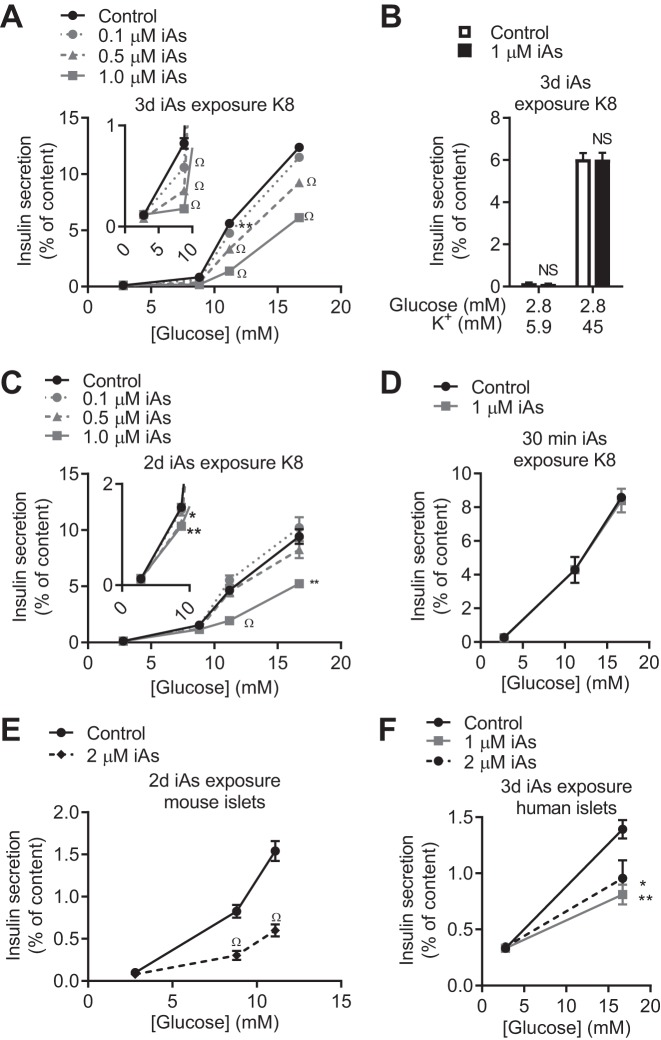
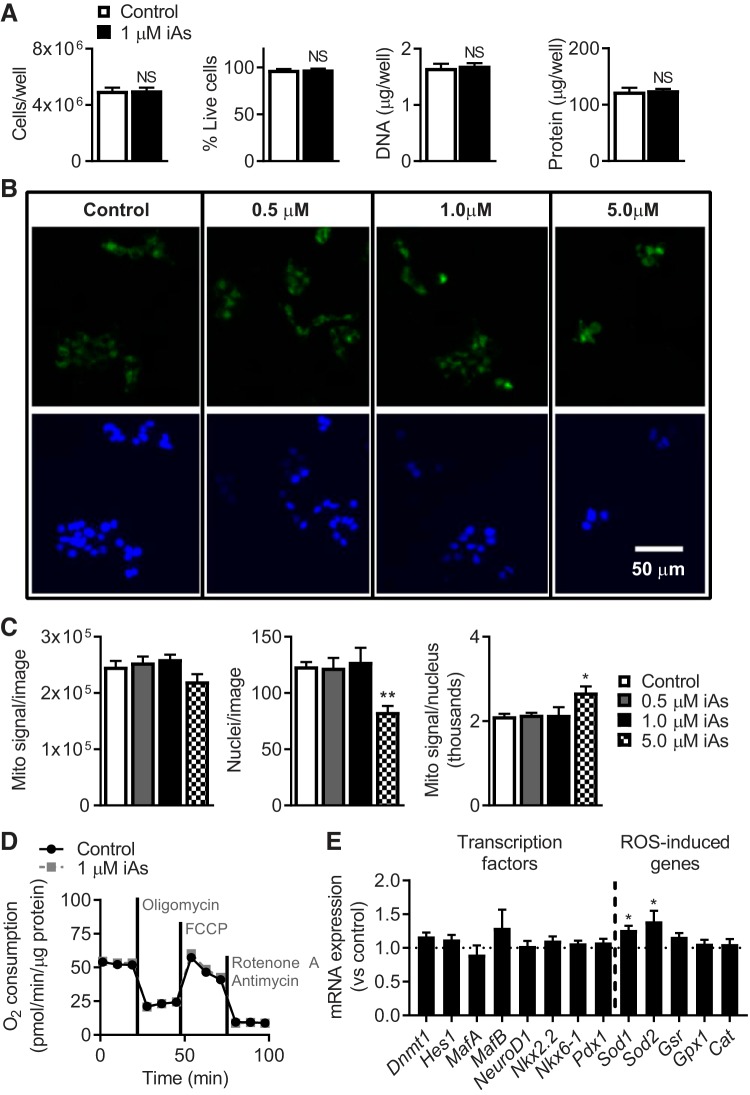
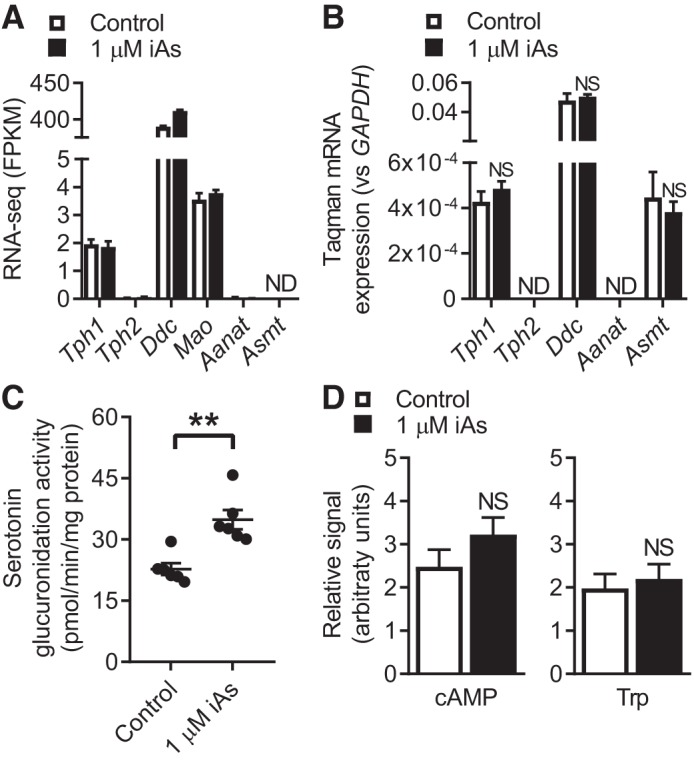
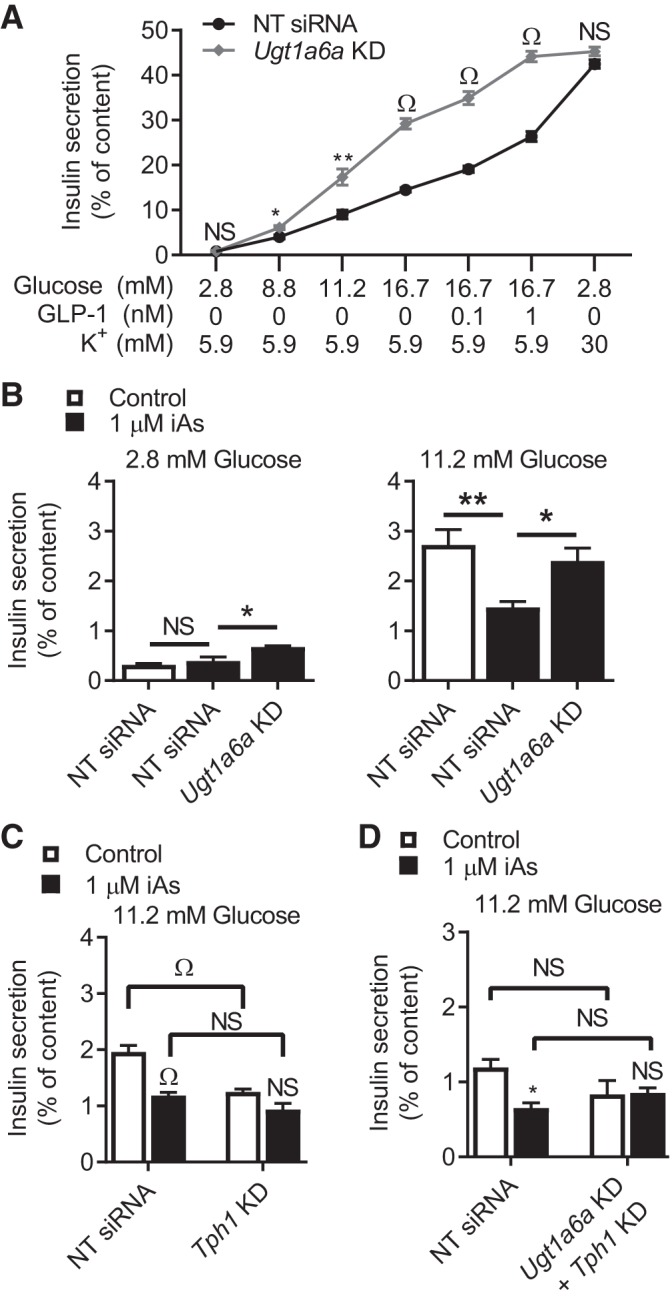
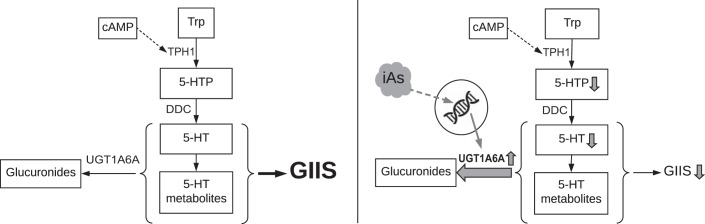
In arsenic-endemic regions of the world, arsenic exposure correlates with diabetes mellitus. Multiple animal models of inorganic arsenic (iAs, as As3+) exposure have revealed that iAs-induced glucose intolerance manifests as a result of pancreatic β-cell dysfunction. To define the mechanisms responsible for this β-cell defect, the MIN6-K8 mouse β-cell line was exposed to environmentally relevant doses of iAs. Exposure to 0.1-1 µM iAs for 3 days significantly decreased glucose-induced insulin secretion (GIIS). Serotonin and its precursor, 5-hydroxytryptophan (5-HTP), were both decreased. Supplementation with 5-HTP, which loads the system with bioavailable 5-HTP and serotonin, rescued GIIS, suggesting that recovery of this pathway was sufficient to restore function. Exposure to iAs was accompanied by an increase in mRNA expression of UDP-glucuronosyltransferase 1 family, polypeptide a6a (Ugt1a6a), a phase-II detoxification enzyme that facilitates the disposal of cyclic amines, including serotonin, via glucuronidation. Elevated Ugt1a6a and UGT1A6 expression levels were observed in mouse and human islets, respectively, following 3 days of iAs exposure. Consistent with this finding, the enzymatic rate of serotonin glucuronidation was increased in iAs-exposed cells. Knockdown by siRNA of Ugt1a6a during iAs exposure restored GIIS in MIN6-K8 cells. This effect was prevented by blockade of serotonin biosynthesis, suggesting that the observed iAs-induced increase in Ugt1a6a affects GIIS by targeting serotonin or serotonin-related metabolites. Although it is not yet clear exactly which element(s) of the serotonin pathway is/are most responsible for iAs-induced GIIS dysfunction, this study provides evidence that UGT1A6A, acting on the serotonin pathway, regulates GIIS under both normal and pathological conditions.
Keywords: arsenic; diabetes; glucuronidation; insulin secretion; serotonin.
Conflict of interest statement
R. M. Sargis has received honoraria from CVS/Health. S. Seino has consulted for JCR Pharmaceuticals and held scientific advisory positions with Kansai Electric Power Medical Research Institute and Servier Laboratories. S. Seino has served on speaker’s bureaus for Novo Nordisk Pharma K.K. and Sumitomo Dainippon Pharma, Co., Ltd., and Novartis Pharma K.K. None of the other authors has any conflicts of interest, financial or otherwise, to disclose.
Figures


References
-
- Argos M, Kalra T, Rathouz PJ, Chen Y, Pierce B, Parvez F, Islam T, Ahmed A, Rakibuz-Zaman M, Hasan R, Sarwar G, Slavkovich V, van Geen A, Graziano J, Ahsan H. Arsenic exposure from drinking water, and all-cause and chronic-disease mortalities in Bangladesh (HEALS): a prospective cohort study. Lancet 376: 252–258, 2010. doi: 10.1016/S0140-6736(10)60481-3. - DOI - PMC - PubMed
-
- Bennet H, Balhuizen A, Medina A, Dekker Nitert M, Ottosson Laakso E, Essén S, Spégel P, Storm P, Krus U, Wierup N, Fex M. Altered serotonin (5-HT) 1D and 2A receptor expression may contribute to defective insulin and glucagon secretion in human type 2 diabetes. Peptides 71: 113–120, 2015. doi: 10.1016/j.peptides.2015.07.008. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials