

Coarse-Grained Protein Dynamics Studies Using Elastic Network Models
- PMID: 30563146
- PMCID: PMC6320916
- DOI: 10.3390/ijms19123899
Coarse-Grained Protein Dynamics Studies Using Elastic Network Models
Abstract
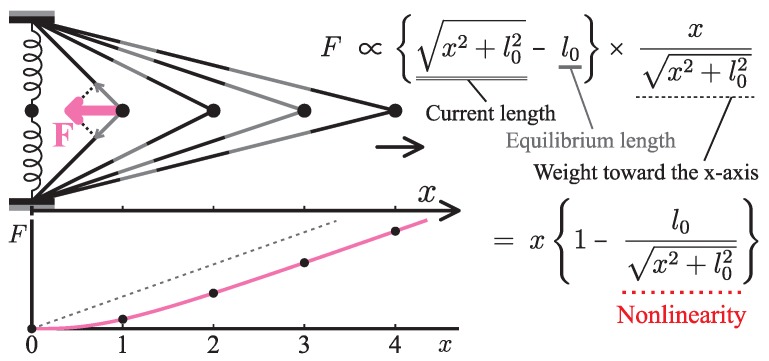
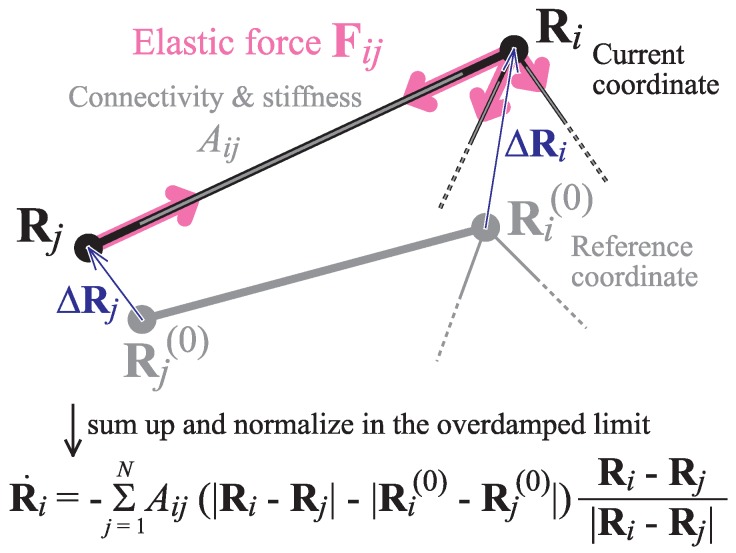
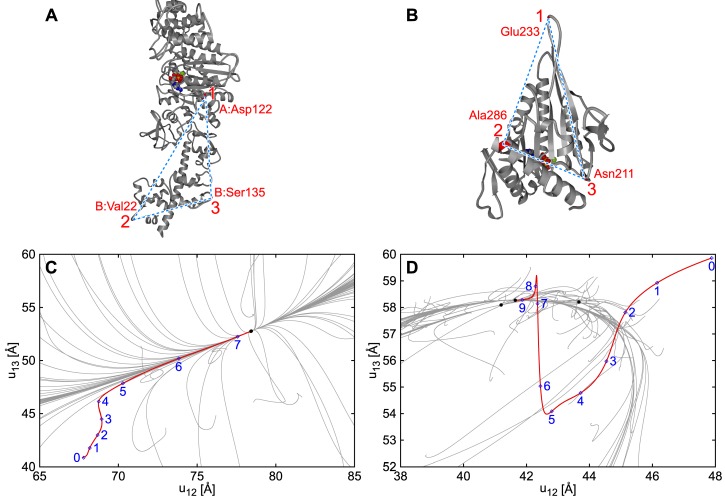
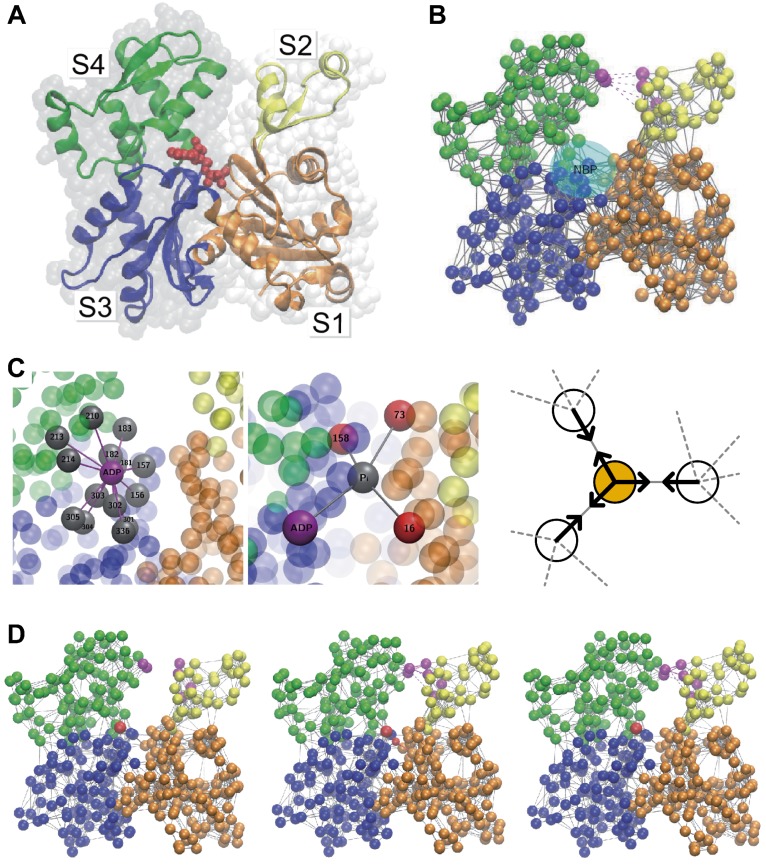
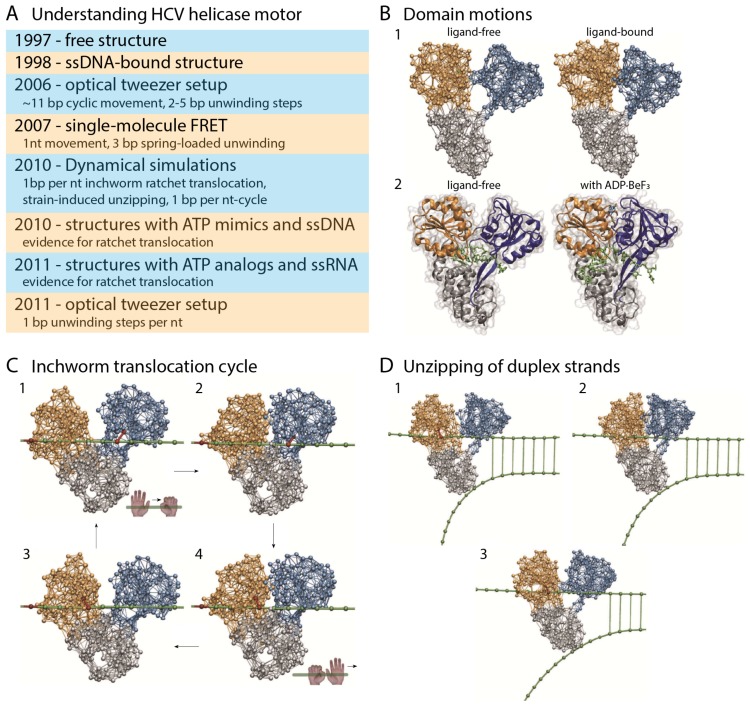
Elastic networks have been used as simple models of proteins to study their slow structural dynamics. They consist of point-like particles connected by linear Hookean springs and hence are convenient for linear normal mode analysis around a given reference structure. Furthermore, dynamic simulations using these models can provide new insights. As the computational cost associated with these models is considerably lower compared to that of all-atom models, they are also convenient for comparative studies between multiple protein structures. In this review, we introduce examples of coarse-grained molecular dynamics studies using elastic network models and their derivatives, focusing on the nonlinear phenomena, and discuss their applicability to large-scale macromolecular assemblies.
Keywords: allostery; coarse-grained model; elastic network; molecular dynamics; molecular machine; nonlinearity; normal mode analysis; protein.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Modeling of Protein Structural Flexibility and Large-Scale Dynamics: Coarse-Grained Simulations and Elastic Network Models.Int J Mol Sci. 2018 Nov 6;19(11):3496. doi: 10.3390/ijms19113496. Int J Mol Sci. 2018. PMID: 30404229 Free PMC article. Review.
-
Inferring a weighted elastic network from partial unfolding with coarse-grained simulations.Proteins. 2014 Jan;82(1):119-29. doi: 10.1002/prot.24381. Epub 2013 Sep 10. Proteins. 2014. PMID: 23900877
-
Efficient prediction of protein conformational pathways based on the hybrid elastic network model.J Mol Graph Model. 2014 Feb;47:25-36. doi: 10.1016/j.jmgm.2013.10.009. Epub 2013 Nov 1. J Mol Graph Model. 2014. PMID: 24296313
-
Modeling Structural Dynamics of Biomolecular Complexes by Coarse-Grained Molecular Simulations.Acc Chem Res. 2015 Dec 15;48(12):3026-35. doi: 10.1021/acs.accounts.5b00338. Epub 2015 Nov 17. Acc Chem Res. 2015. PMID: 26575522 Review.
-
Multiscale design of coarse-grained elastic network-based potentials for the μ opioid receptor.J Mol Model. 2016 Sep;22(9):227. doi: 10.1007/s00894-016-3092-z. Epub 2016 Aug 26. J Mol Model. 2016. PMID: 27566318
Cited by
-
Explainable machine learning for predicting distant metastases in renal cell carcinoma patients: a population-based retrospective study.Front Med (Lausanne). 2025 Jul 29;12:1624198. doi: 10.3389/fmed.2025.1624198. eCollection 2025. Front Med (Lausanne). 2025. PMID: 40800127 Free PMC article.
-
In silico investigation and surmounting of Lipopolysaccharide barrier in Gram-Negative Bacteria: How far has molecular dynamics Come?Comput Struct Biotechnol J. 2022 Oct 29;20:5886-5901. doi: 10.1016/j.csbj.2022.10.039. eCollection 2022. Comput Struct Biotechnol J. 2022. PMID: 36382192 Free PMC article. Review.
-
Revolutionizing Molecular Design for Innovative Therapeutic Applications through Artificial Intelligence.Molecules. 2024 Sep 29;29(19):4626. doi: 10.3390/molecules29194626. Molecules. 2024. PMID: 39407556 Free PMC article. Review.
-
Protein Function Analysis through Machine Learning.Biomolecules. 2022 Sep 6;12(9):1246. doi: 10.3390/biom12091246. Biomolecules. 2022. PMID: 36139085 Free PMC article. Review.
-
"Dividing and Conquering" and "Caching" in Molecular Modeling.Int J Mol Sci. 2021 May 10;22(9):5053. doi: 10.3390/ijms22095053. Int J Mol Sci. 2021. PMID: 34068835 Free PMC article. Review.
References
-
- Shaw D.E., Dror R.O., Salmon J.K., Grossman J.P., Mackenzie K.M., Bank J.A., Young C., Deneroff M.M., Batson B., Bowers K.J., et al. Millisecond-scale molecular dynamics simulations on Anton; Proceedings of the Conference on High Performance Computing, Networking, Storage and Analysis (SC09); Portland, OR, USA. 14–20 November 2009.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
