

Evolution of Plant Architecture, Functional Diversification and Divergent Evolution in the Genus Atractocarpus (Rubiaceae) for New Caledonia
- PMID: 30564258
- PMCID: PMC6288547
- DOI: 10.3389/fpls.2018.01775
Evolution of Plant Architecture, Functional Diversification and Divergent Evolution in the Genus Atractocarpus (Rubiaceae) for New Caledonia
Abstract
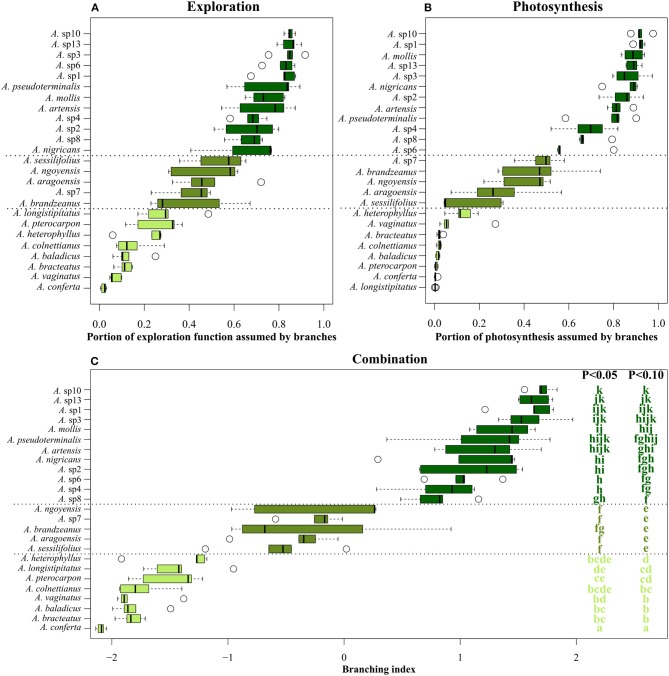
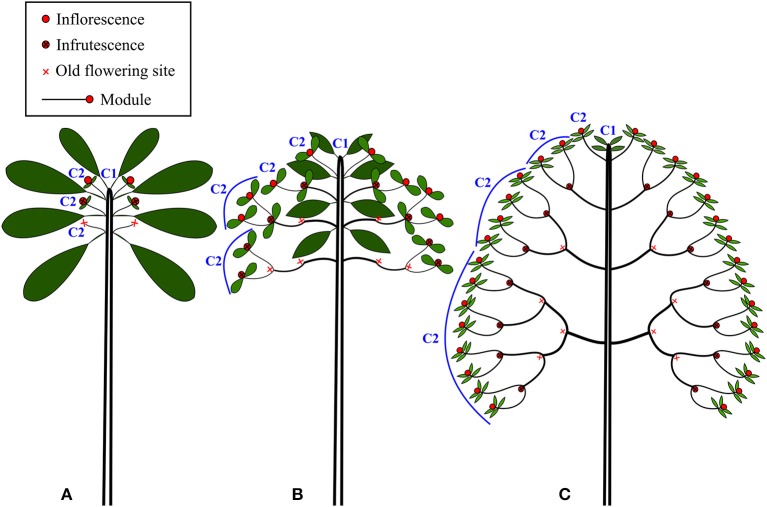
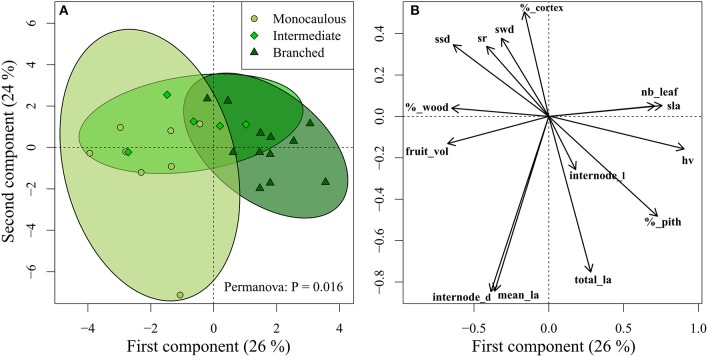
The diversification of ecological roles and related adaptations in closely related species within a lineage is one of the most important processes linking plant evolution and ecology. Plant architecture offers a robust framework to study these processes as it can highlight how plant structure influences plant diversification and ecological strategies. We investigated a case of gradual evolution of branching architecture in Atractocarpus spp. (Rubiaceae), forming a monophyletic group in New Caledonia that has diversified rapidly, predominantly in rainforest understory habitats. We used a transdisciplinary approach to depict architectural variations and revealed multiple evolutionary transitions from a branched (Stone's architectural model) to a monocaulous habit (Corner's architectural model), which involved the functional reduction of branches into inflorescences. We propose an integrative functional index that assesses branching incidence on functional traits influencing both assimilation and exploration functions. We showed that architectural transitions correlate with ecologically important functional traits. Variation in ecologically important traits among closely relatives, as supported by the architectural analysis, is suggestive of intense competition that favored divergence among locally coexisting species. We propose that Pleistocene climatic fluctuations causing expansion and contraction of rainforest could also have offered ecological opportunities for colonizers in addition to the process of divergent evolution.
Keywords: Island; branching index; convergence; corner's rules; gardenieae; rainforest; treelet; understory.
Figures



References
-
- Ackerly D. D. (1996). Canopy structure and dynamics: integration of growth processes in tropical pioneer trees, in Tropical Forest Plant Ecophysiology, eds Mulkey S. S., Chazdon R. L., Smith A. P. (New York, NY: Chapman & Hall; ), 619–658.
-
- Ackerly D. D., Donoghue M. J. (1998). Leaf size, sapling allometry, and corner's rules: phylogeny and correlated evolution in maples (Acer). Am. Nat. 152, 767–791. - PubMed
-
- Alberch P., Gould S. J., Oster G. F., Wake D. B. (1979). Size and shape in ontogeny and phylogeny. Paleobiology 5, 296–317. 10.1017/S0094837300006588 - DOI
LinkOut - more resources
Full Text Sources
Miscellaneous