

Circulating AQP4-specific auto-antibodies alone can induce neuromyelitis optica spectrum disorder in the rat
- PMID: 30564980
- PMCID: PMC6514074
- DOI: 10.1007/s00401-018-1950-8
Circulating AQP4-specific auto-antibodies alone can induce neuromyelitis optica spectrum disorder in the rat
Abstract
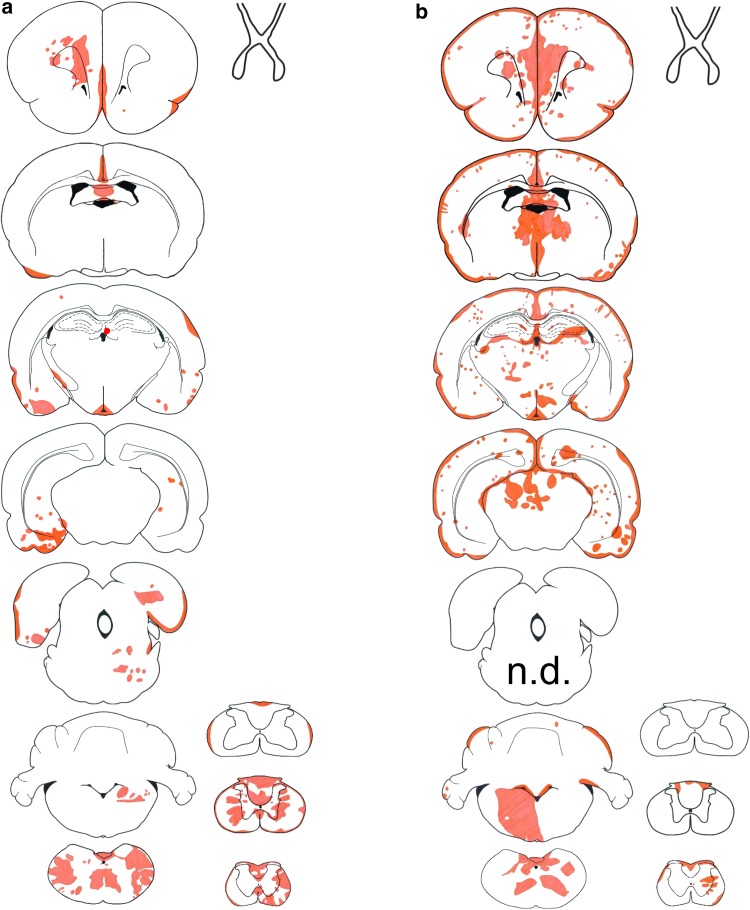
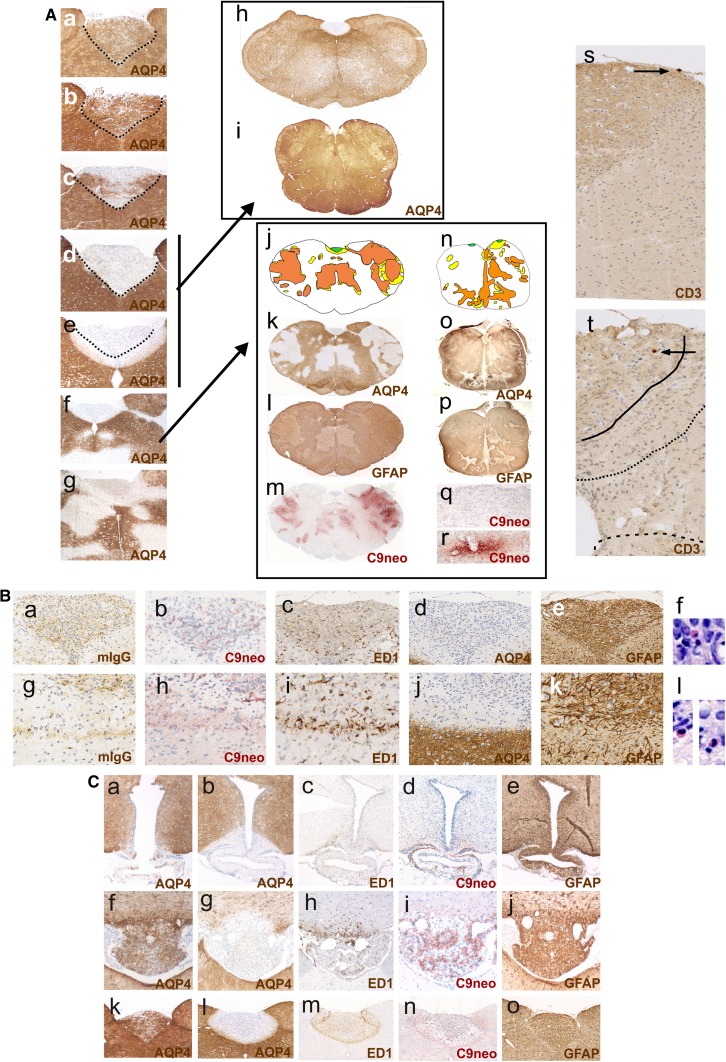
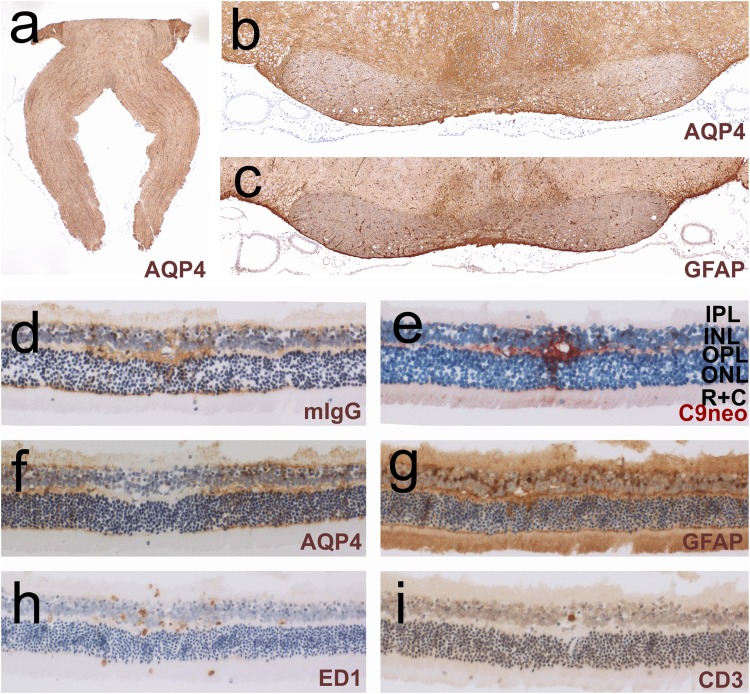
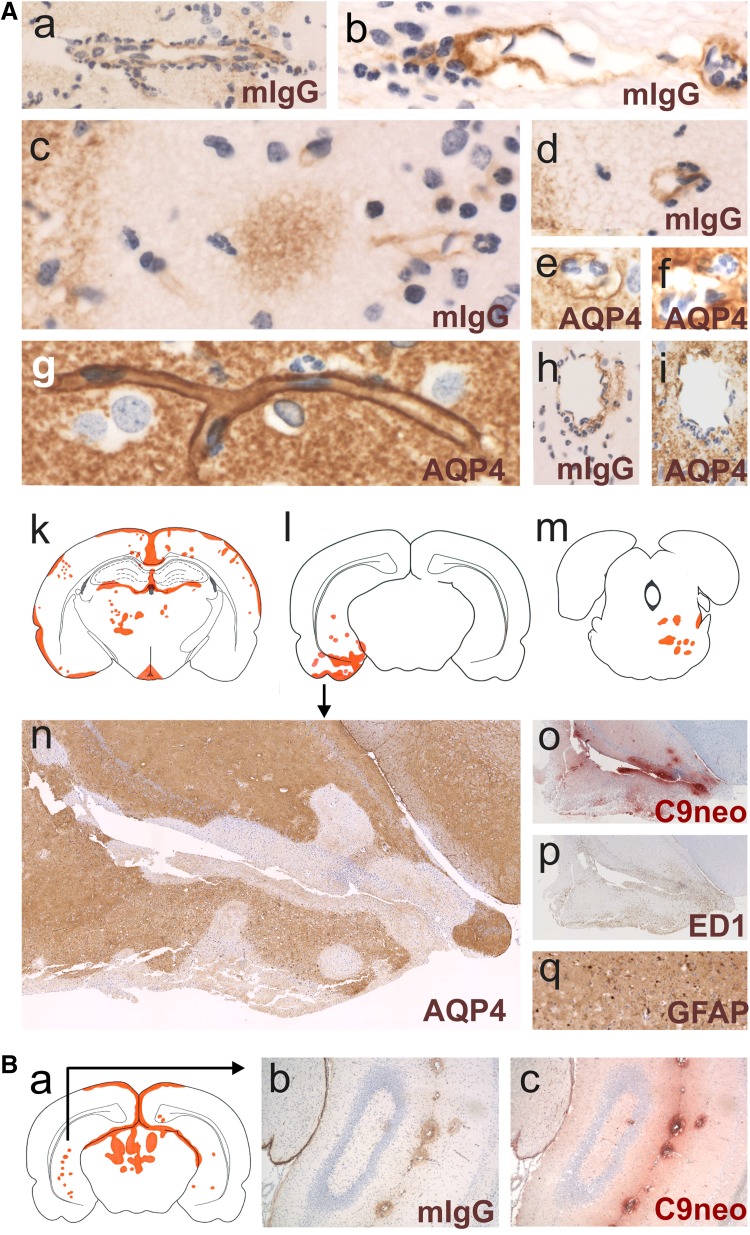
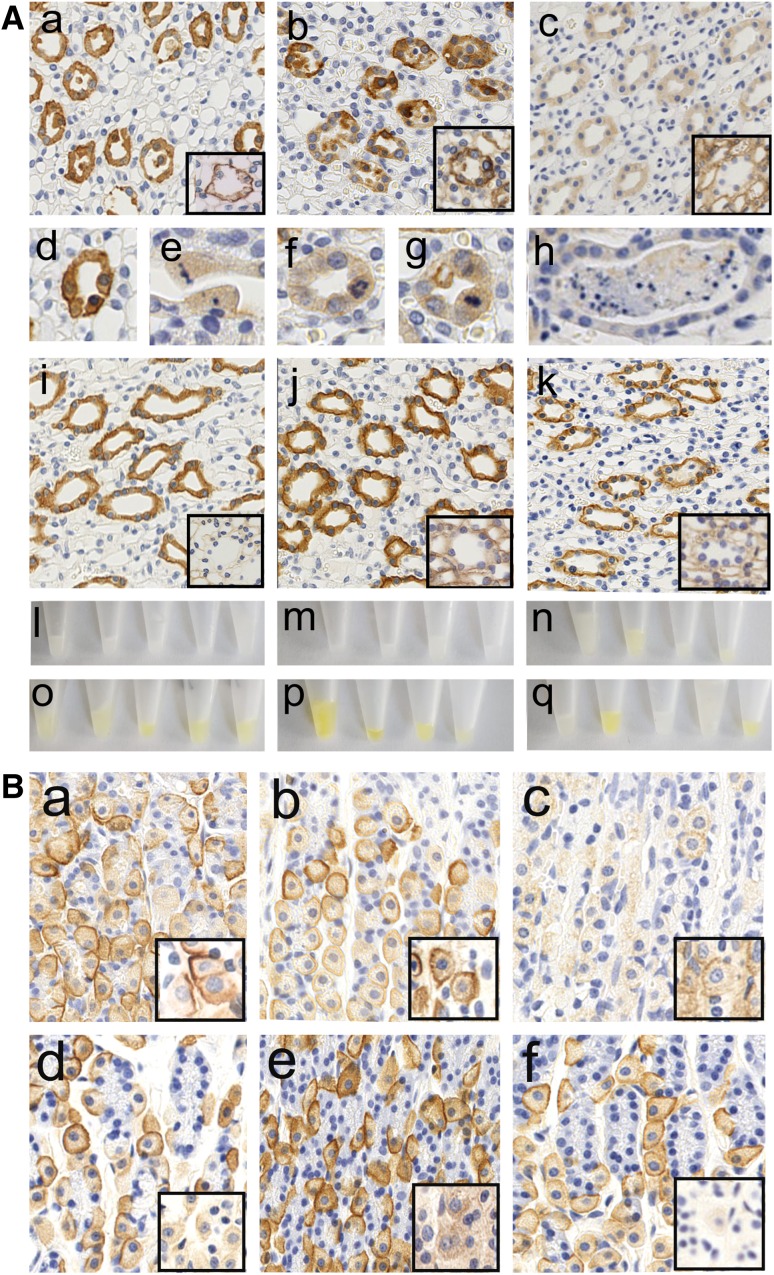
It is well established that the binding of pathogenic aquaporin-4 (AQP4)-specific autoantibodies to astrocytes may initiate a cascade of events culminating in the destruction of these cells and in the formation of large tissue-destructive lesions typical for patients with neuromyelitis optica spectrum disorders (NMOSD). To date, not a single experimental study has shown that the systemic presence of the antibody alone can induce any damage to the central nervous system (CNS), while pathological studies on brains of NMOSD patients suggested that there might be ways for antibody entry and subsequent tissue damage. Here, we systemically applied a highly pathogenic, monoclonal antibody with high affinity to AQP4 over prolonged period of time to rats, and show that AQP4-abs can enter the CNS on their own, via circumventricular organs and meningeal or parenchymal blood vessels, that these antibodies initiate the formation of radically different lesions with AQP4 loss, depending on their mode and site of entry, and that lesion formation is much more efficient in the presence of encephalitogenic T-cell responses. We further demonstrate that the established tissue-destructive lesions trigger the formation of additional lesions by short and far reaching effects on blood vessels and their branches, and that AQP4-abs have profound effects on the AQP4 expression in peripheral tissues which counter-act possible titer loss by antibody absorption outside the CNS. Cumulatively, these data indicate that directly induced pathological changes caused by AQP4-abs inside and outside the CNS are efficient drivers of disease evolution in seropositive organisms.
Keywords: Aquaporin-4; Aquaporin-4-specific antibodies; Kidney; Lesions; Neuromyelitis optica; T cells.
Conflict of interest statement
SH, KS, MN, IT, DB, PP, YT, IN, TM, and MB: the authors declare that they have no conflict of interest. HL received honoraria for lectures from Novartis, Biogen, and Sanofi Aventis. Moreover, he is a member of advisory boards at Roche and Medday. The University Hospital and Medical University of Innsbruck (Austria, MR) receives payments for antibody assays (MOG, AQP4, and other autoantibodies) and for MOG and AQP4 antibody validation experiments organized by Euroimmun (Lübeck, Germany). K. Fujihara serves on scientific advisory boards for Bayer, Biogen, Mitsubishi Tanabe, Novartis, Chugai, Ono, Nihon, Merck Serono, Alexion, and MedImmune; has received funding for travel and speaker honoraria from Bayer, Biogen, Eisai, Mitsubishi Tanabe, Novartis, Astellas, Takeda, Asahi Kasei Medical, Daiichi-Sankyo, and Nihon; serves as an editorial board member of Clinical and Experimental Neuroimmunology, Frontiers in Neurology (section Multiple Sclerosis and Neuroimmunology) and Neurology: Neuroimmunology and Neuroinflammation, and an advisory board member of Sri Lanka Journal of Neurology; has received research support from Bayer, Biogen, Asahi Kasei Medical, The Chemo-Sero-Therapeutic Research Institute, Teva, Mitsubishi Tanabe, Teijin, Chugai, Ono, Nihon, and Genzyme; is funded by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan and by Grants-in-Aid for Scientific Research from the Ministry of Health, Welfare and Labor of Japan.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
