

AMPK signaling linked to the schizophrenia-associated 1q21.1 deletion is required for neuronal and sleep maintenance
- PMID: 30566533
- PMCID: PMC6317821
- DOI: 10.1371/journal.pgen.1007623
AMPK signaling linked to the schizophrenia-associated 1q21.1 deletion is required for neuronal and sleep maintenance
Abstract
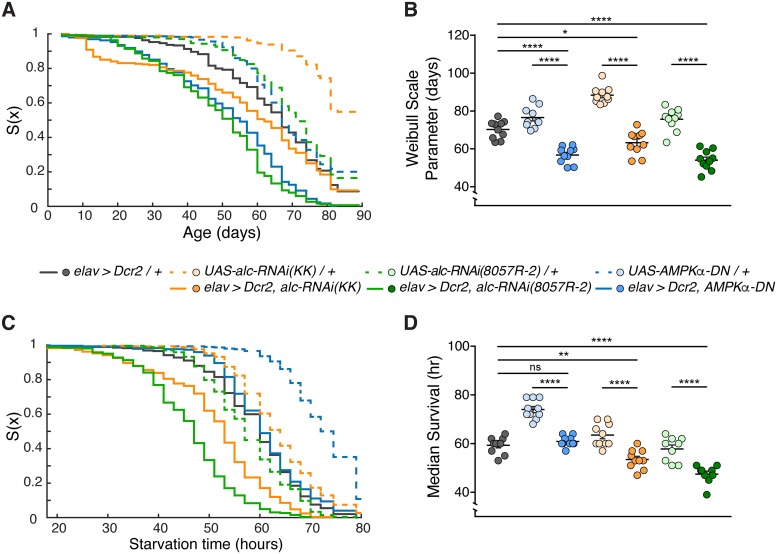
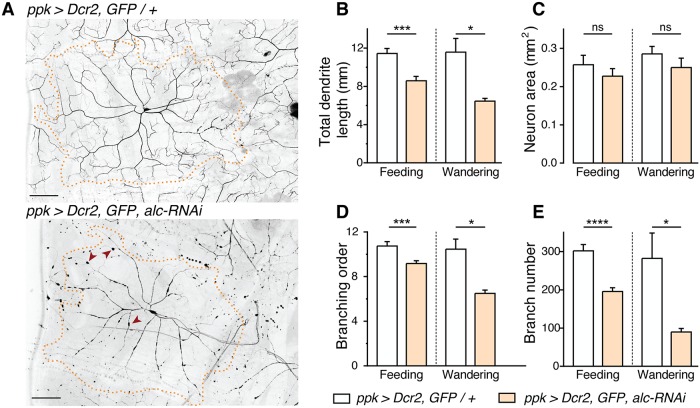
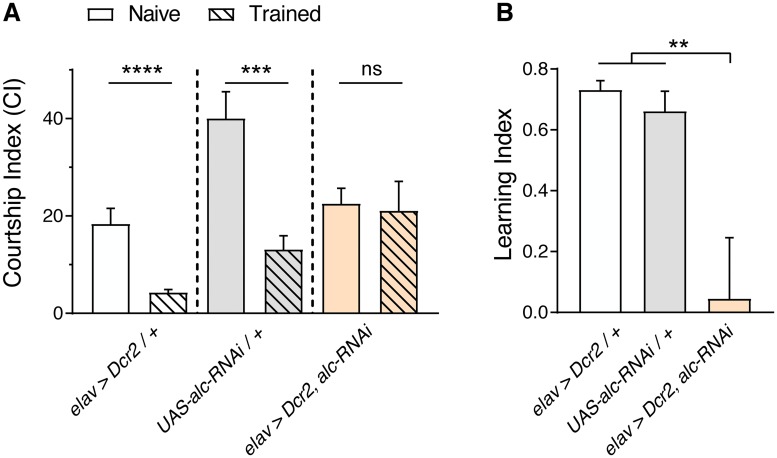
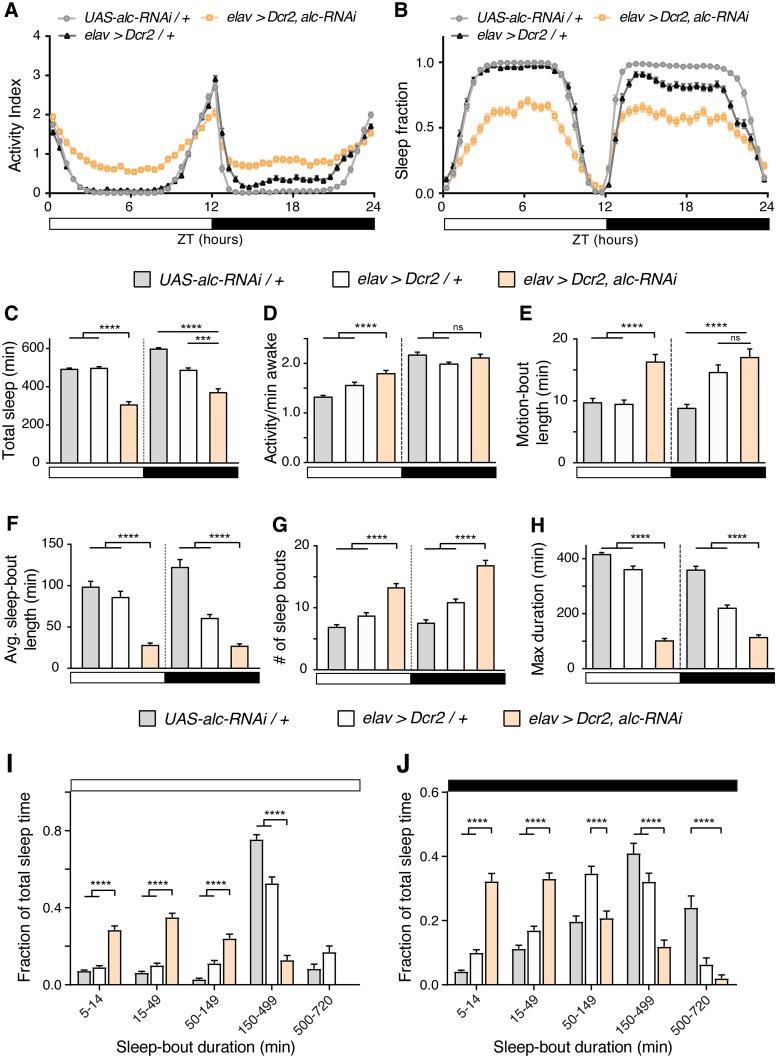
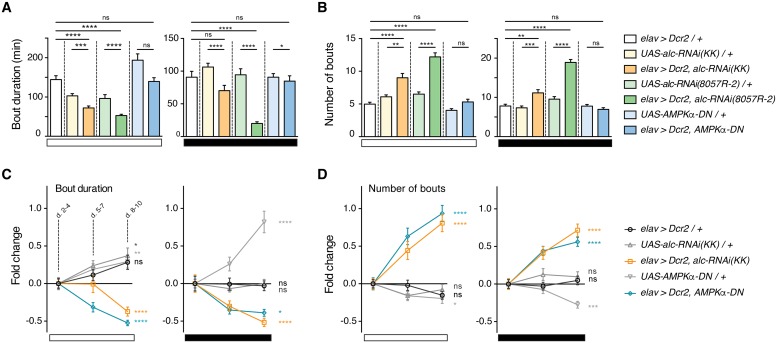
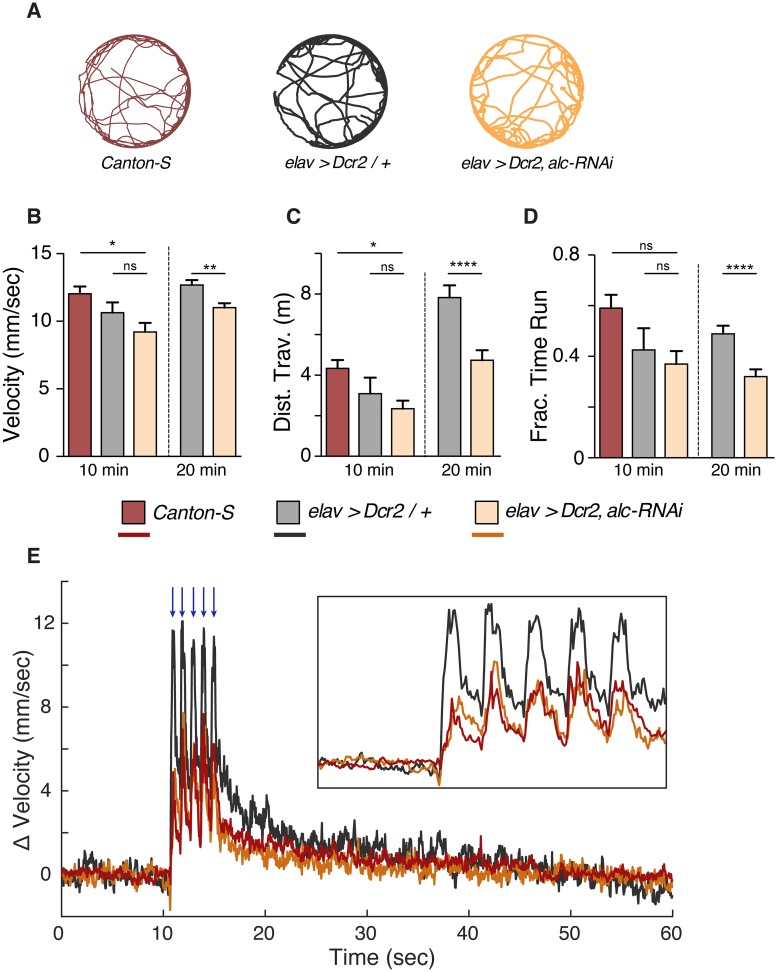
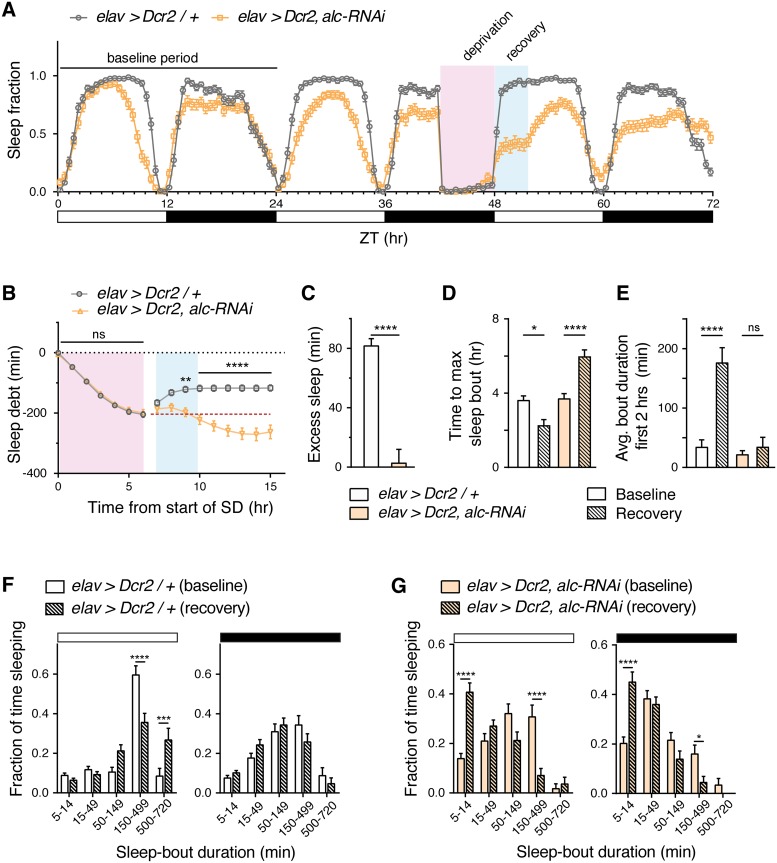
The human 1q21.1 deletion of ten genes is associated with increased risk of schizophrenia. This deletion involves the β-subunit of the AMP-activated protein kinase (AMPK) complex, a key energy sensor in the cell. Although neurons have a high demand for energy and low capacity to store nutrients, the role of AMPK in neuronal physiology is poorly defined. Here we show that AMPK is important in the nervous system for maintaining neuronal integrity and for stress survival and longevity in Drosophila. To understand the impact of this signaling system on behavior and its potential contribution to the 1q21.1 deletion syndrome, we focused on sleep, an important role of which is proposed to be the reestablishment of neuronal energy levels that are diminished during energy-demanding wakefulness. Sleep disturbances are one of the most common problems affecting individuals with psychiatric disorders. We show that AMPK is required for maintenance of proper sleep architecture and for sleep recovery following sleep deprivation. Neuronal AMPKβ loss specifically leads to sleep fragmentation and causes dysregulation of genes believed to play a role in sleep homeostasis. Our data also suggest that AMPKβ loss may contribute to the increased risk of developing mental disorders and sleep disturbances associated with the human 1q21.1 deletion.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Fejgin K, Nielsen J, Birknow MR, Bastlund JF, Nielsen V, Lauridsen JB, et al. A mouse model that recapitulates cardinal features of the 15q13.3 microdeletion syndrome including schizophrenia- and epilepsy-related alterations. Biol Psychiatry. 2014;76(2):128–37. 10.1016/j.biopsych.2013.08.014 . - DOI - PubMed
-
- Ingason A, Giegling I, Cichon S, Hansen T, Rasmussen HB, Nielsen J, et al. A large replication study and meta-analysis in European samples provides further support for association of AHI1 markers with schizophrenia. Hum Mol Genet. 2010;19(7):1379–86. Epub 2010/01/15. 10.1093/hmg/ddq009 . - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
