

FAD-sequestering proteins protect mycobacteria against hypoxic and oxidative stress
- PMID: 30567740
- PMCID: PMC6393599
- DOI: 10.1074/jbc.RA118.006237
FAD-sequestering proteins protect mycobacteria against hypoxic and oxidative stress
Abstract
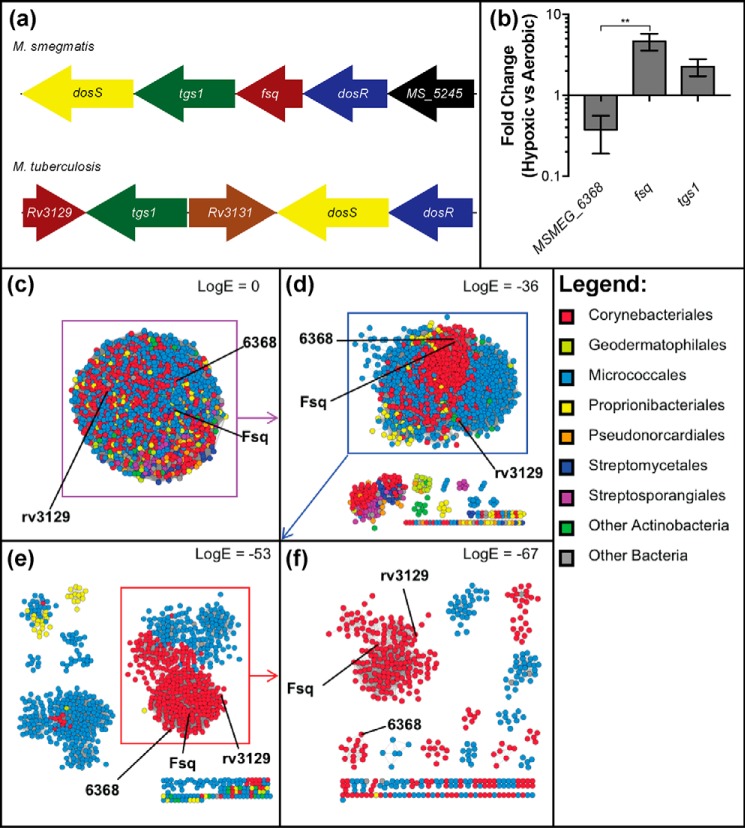
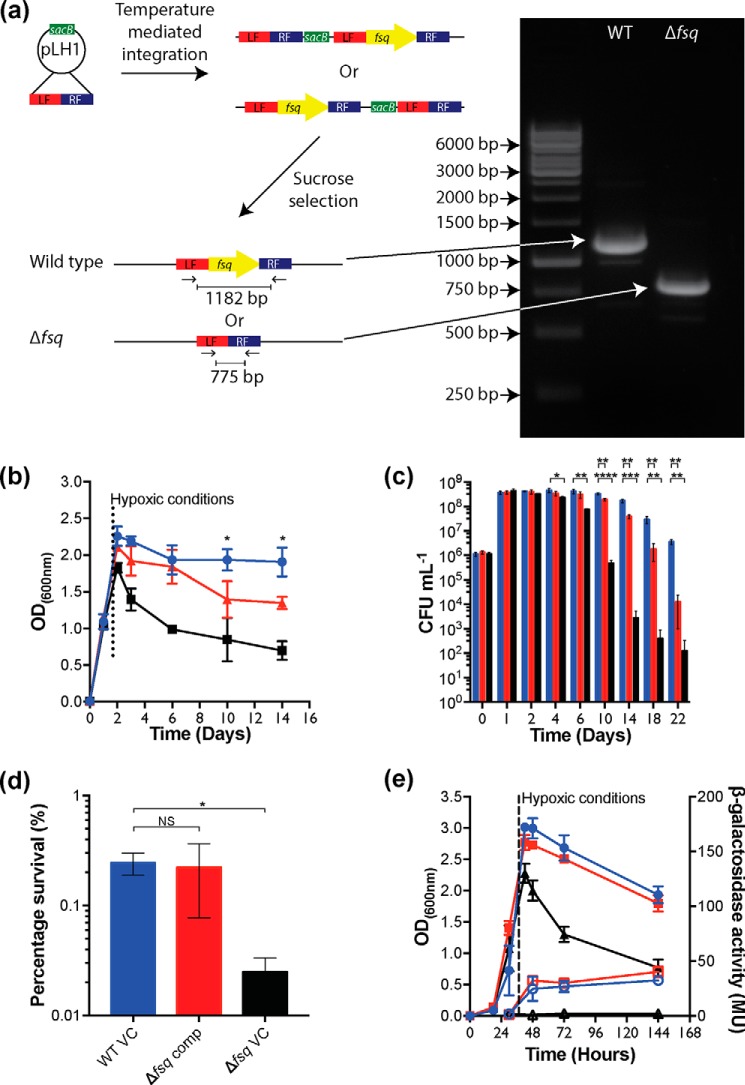
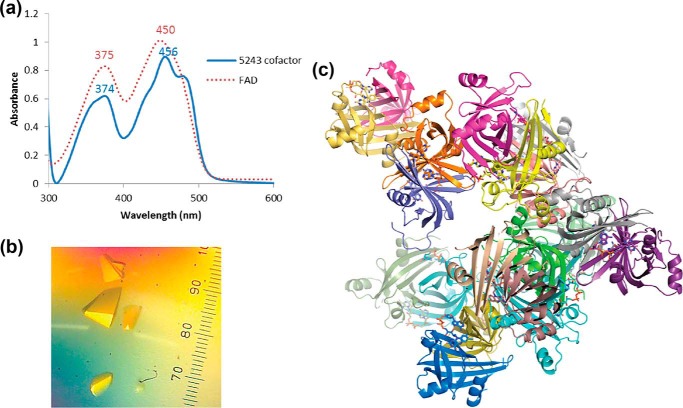
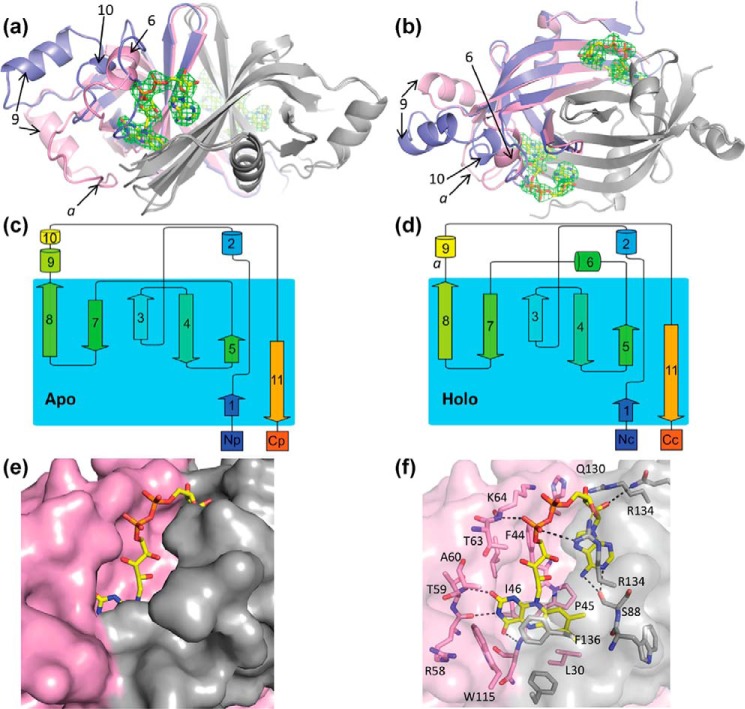
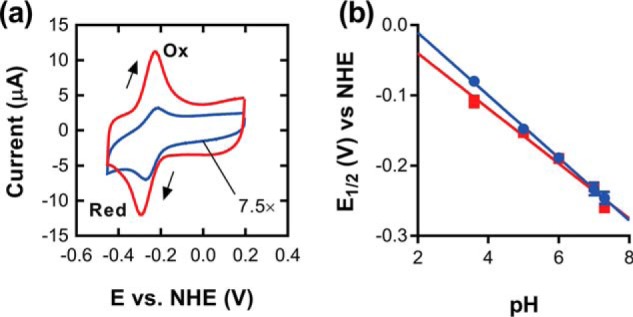
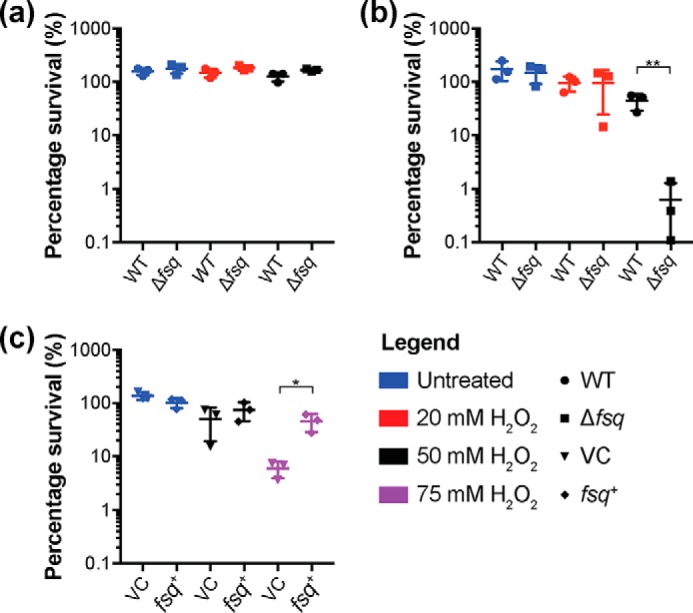
The ability to persist in the absence of growth triggered by low oxygen levels is a critical process for the survival of mycobacterial species in many environmental niches. MSMEG_5243 (fsq), a gene of unknown function in Mycobacterium smegmatis, is up-regulated in response to hypoxia and regulated by DosRDosS/DosT, an oxygen- and redox-sensing two-component system that is highly conserved in mycobacteria. In this communication, we demonstrate that MSMEG_5243 is a
Keywords: reactive oxygen species (ROS); mycobacteria; Mycobacterium smegmatis; hypoxia; flavin; flavin adenine dinucleotide (FAD); bacterial genetics; oxidative stress; anoxia; dos regulon; DosR; oxidoreductase.
© 2019 Harold et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Massey V. (1994) Activation of molecular-oxygen by flavins and flavoproteins. J. Biol. Chem. 269, 22459–22462 - PubMed
-
- Piddington D. L., Fang F. C., Laessig T., Cooper A. M., Orme I. M., and Buchmeier N. A. (2001) Cu,Zn superoxide dismutase of Mycobacterium tuberculosis contributes to survival in activated macrophages that are generating an oxidative burst. Infect. Immun. 69, 4980–4987 10.1128/IAI.69.8.4980-4987.2001 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
