

Human kidney on a chip assessment of polymyxin antibiotic nephrotoxicity
- PMID: 30568031
- PMCID: PMC6338315
- DOI: 10.1172/jci.insight.123673
Human kidney on a chip assessment of polymyxin antibiotic nephrotoxicity
Abstract
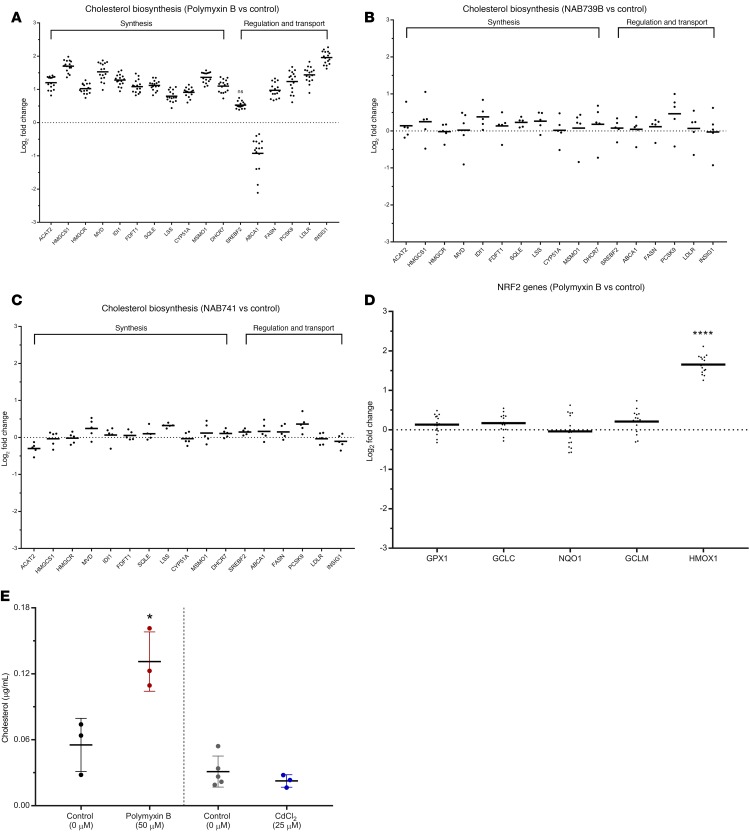
Drug-induced kidney injury, largely caused by proximal tubular intoxicants, limits development and clinical use of new and approved drugs. Assessing preclinical nephrotoxicity relies on animal models that are frequently insensitive; thus, potentially novel techniques - including human microphysiological systems, or "organs on chips" - are proposed to accelerate drug development and predict safety. Polymyxins are potent antibiotics against multidrug-resistant microorganisms; however, clinical use remains restricted because of high risk of nephrotoxicity and limited understanding of toxicological mechanisms. To mitigate risks, structural analogs of polymyxins (NAB739 and NAB741) are currently in clinical development. Using a microphysiological system to model human kidney proximal tubule, we exposed cells to polymyxin B (PMB) and observed significant increases of injury signals, including kidney injury molecule-1 KIM-1and a panel of injury-associated miRNAs (each P < 0.001). Surprisingly, transcriptional profiling identified cholesterol biosynthesis as the primary cellular pathway induced by PMB (P = 1.22 ×10-16), and effluent cholesterol concentrations were significantly increased after exposure (P < 0.01). Additionally, we observed no upregulation of the nuclear factor (erythroid derived-2)-like 2 pathway, despite this being a common pathway upregulated in response to proximal tubule toxicants. In contrast with PMB exposure, minimal changes in gene expression, injury biomarkers, and cholesterol concentrations were observed in response to NAB739 and NAB741. Our findings demonstrate the preclinical safety of NAB739 and NAB741 and reveal cholesterol biosynthesis as a potentially novel pathway for PMB-induced injury. To our knowledge, this is the first demonstration of a human-on-chip platform used for simultaneous safety testing of new chemical entities and defining unique toxicological pathway responses of an FDA-approved molecule.
Keywords: Cell stress; Nephrology; Toxicology; Toxins/drugs/xenobiotics.
Conflict of interest statement
Figures
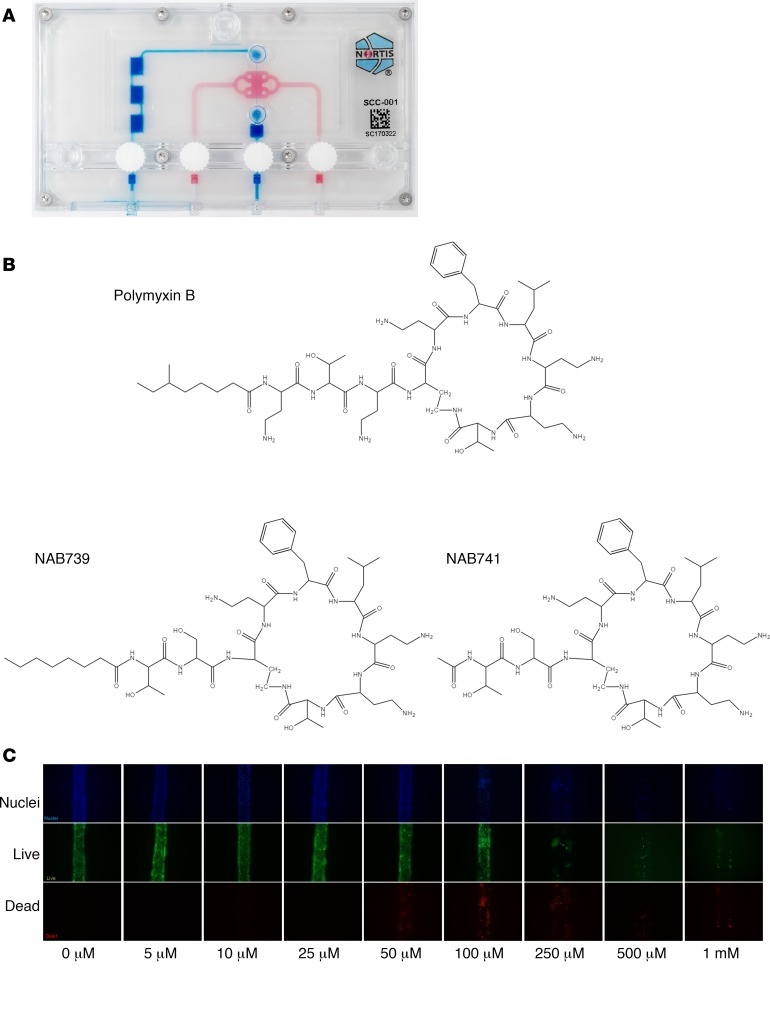
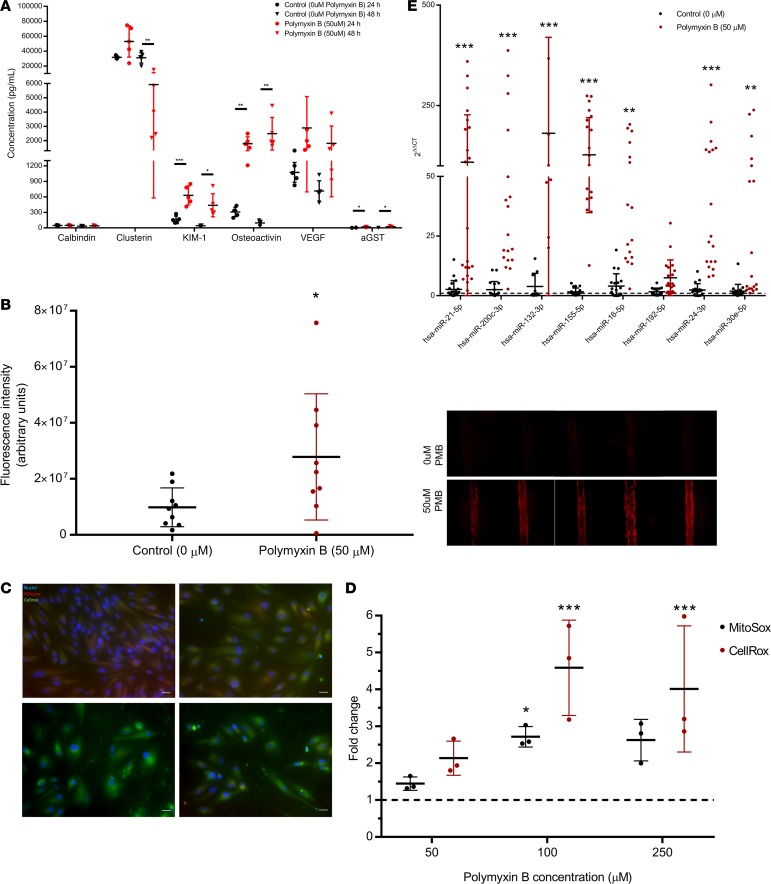
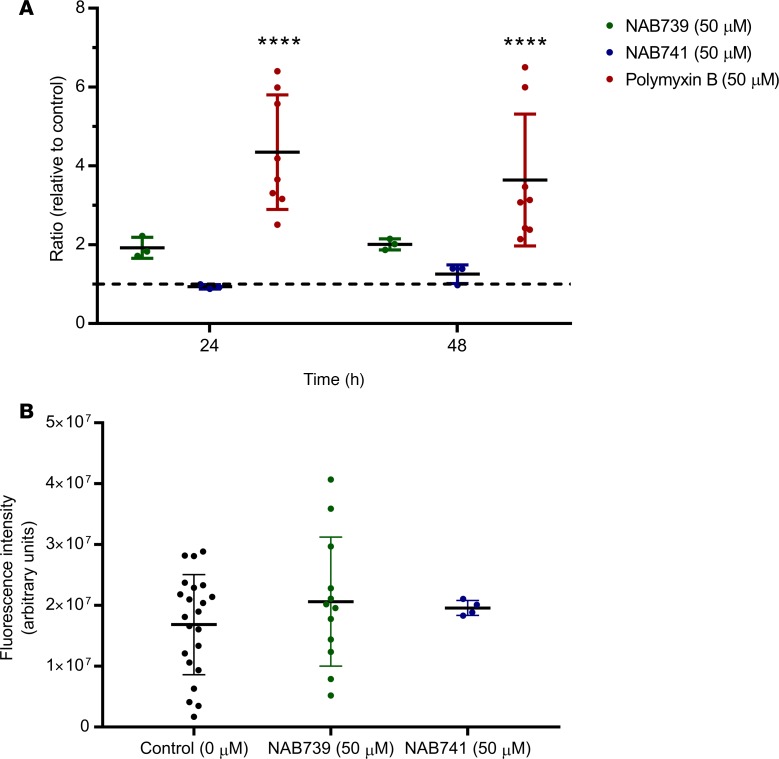
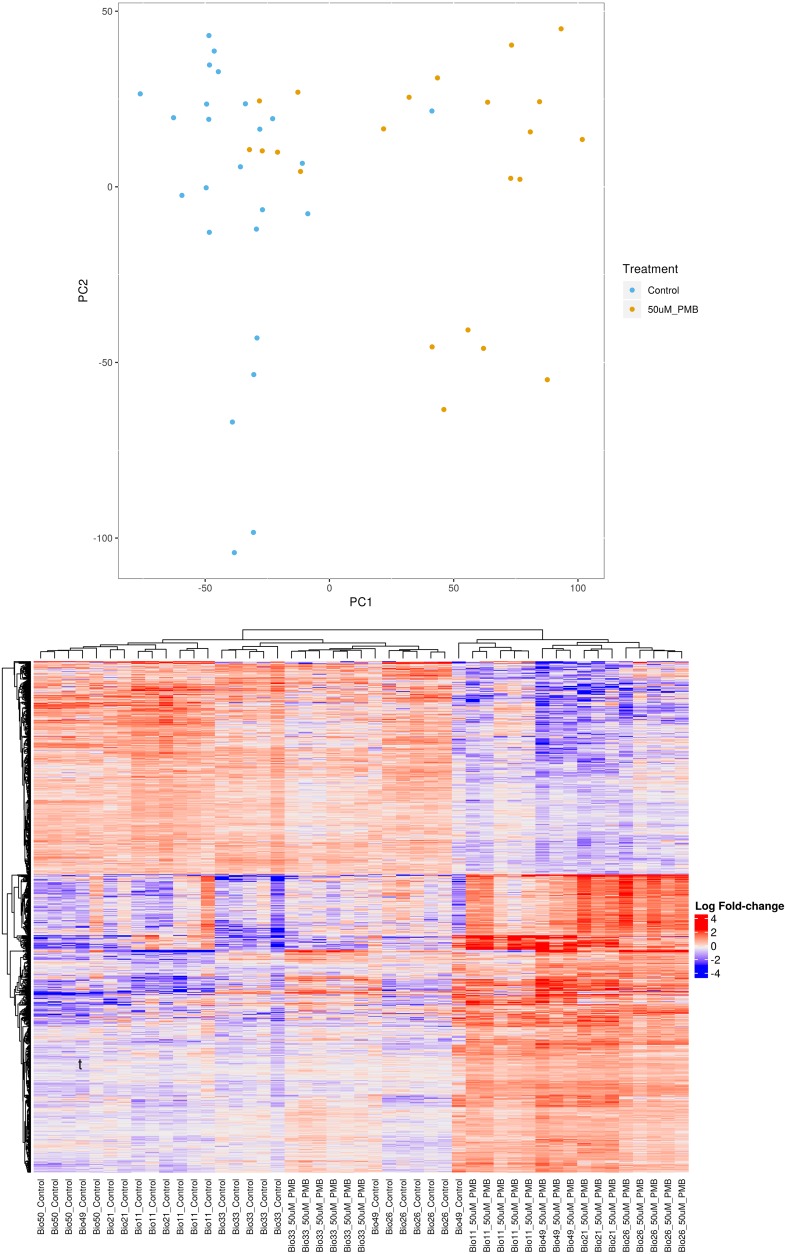
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
