

OsDIRP1, a Putative RING E3 Ligase, Plays an Opposite Role in Drought and Cold Stress Responses as a Negative and Positive Factor, Respectively, in Rice (Oryza sativa L.)
- PMID: 30568669
- PMCID: PMC6290360
- DOI: 10.3389/fpls.2018.01797
OsDIRP1, a Putative RING E3 Ligase, Plays an Opposite Role in Drought and Cold Stress Responses as a Negative and Positive Factor, Respectively, in Rice (Oryza sativa L.)
Abstract
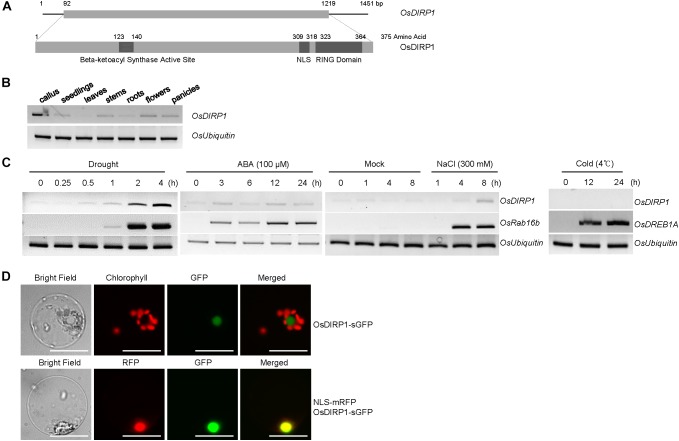
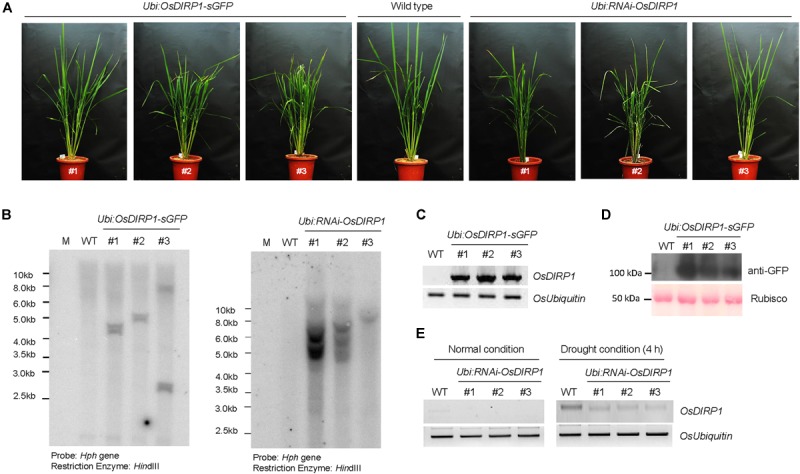
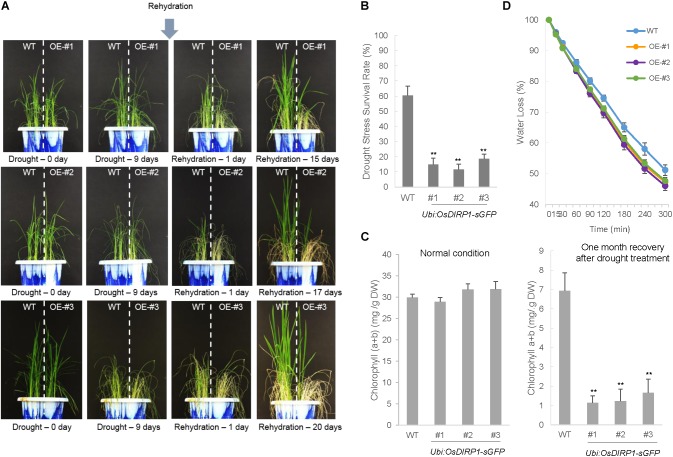
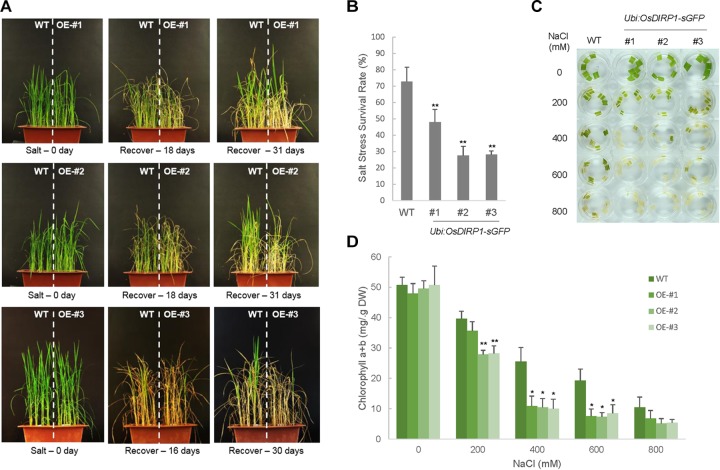
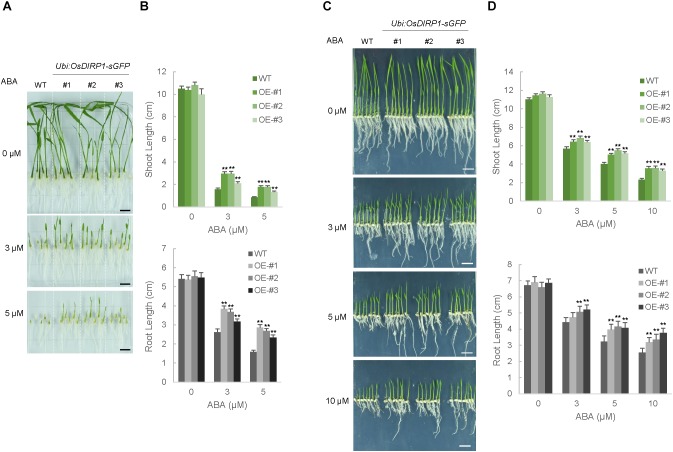
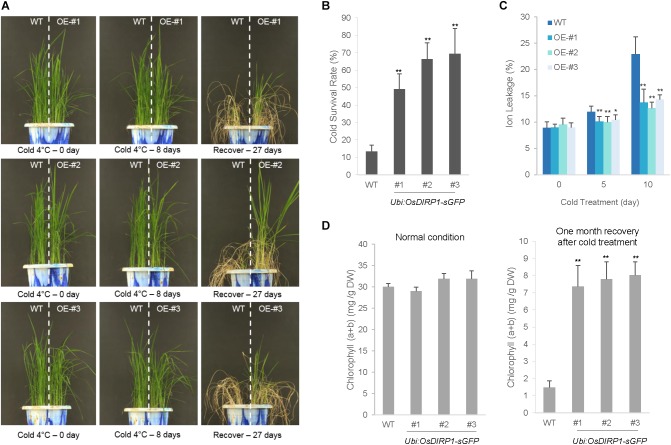
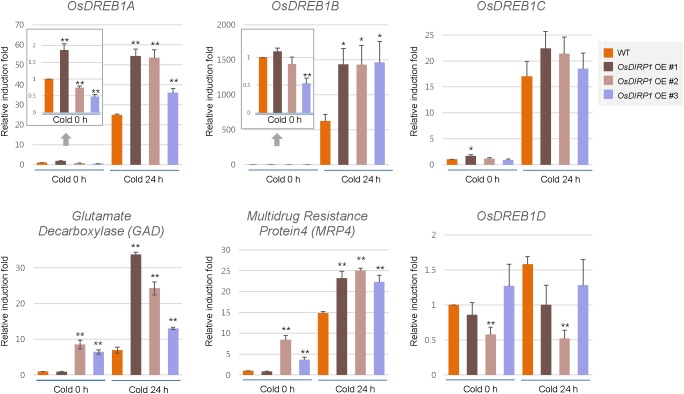
As higher plants are sessile organisms, they are unable to move to more favorable places; thus, they have developed the ability to survive under potentially detrimental conditions. Ubiquitination is a crucial post-translational protein modification and participates in abiotic stress responses in higher plants. In this study, we identified and characterized OsDIRP1 (Oryza sativa Drought-Induced RING Protein 1), a nuclear-localized putative RING E3 ubiquitin (Ub) ligase in rice (Oryza sativa L.). OsDIRP1 expression was induced by drought, high salinity, and abscisic acid (ABA) treatment, but not by low temperature (4°C) stress, suggesting that OsDIRP1 is differentially regulated by different abiotic stresses. To investigate its possible role in abiotic stress responses, OsDIRP1-overexpressing transgenic rice plants (Ubi:OsDIRP1-sGFP) were generated, and their phenotypes were analyzed. The T4 Ubi:OsDIRP1-sGFP lines showed decreased tolerance to drought and salt stress as compared to wild-type rice plants. Moreover, Ubi:OsDIRP1-sGFP progeny were less sensitive to ABA than the wild-type during both germination and post-germination growth. In contrast, Ubi:OsDIRP1-sGFP plants exhibited markedly higher tolerance to prolonged cold (4°C) treatment. These results suggest that OsDIRP1 acts as a negative regulator during drought and salt stress, whereas it functions as a positive factor during the cold stress response in rice.
Keywords: RING E3 Ub ligase; abscisic acid; cold stress; drought stress; opposite response; rice (Oryza sativa).
Figures
Similar articles
-
CaPUB1, a Hot Pepper U-box E3 Ubiquitin Ligase, Confers Enhanced Cold Stress Tolerance and Decreased Drought Stress Tolerance in Transgenic Rice (Oryza sativa L.).Mol Cells. 2016 Mar;39(3):250-7. doi: 10.14348/molcells.2016.2290. Epub 2015 Dec 16. Mol Cells. 2016. PMID: 26674966 Free PMC article.
-
Oryza sativa drought-, heat-, and salt-induced RING finger protein 1 (OsDHSRP1) negatively regulates abiotic stress-responsive gene expression.Plant Mol Biol. 2020 Jun;103(3):235-252. doi: 10.1007/s11103-020-00989-x. Epub 2020 Mar 21. Plant Mol Biol. 2020. PMID: 32206999
-
The Arabidopsis RING E3 ubiquitin ligase AtAIRP2 plays combinatory roles with AtAIRP1 in abscisic acid-mediated drought stress responses.Plant Physiol. 2011 Dec;157(4):2240-57. doi: 10.1104/pp.111.185595. Epub 2011 Oct 10. Plant Physiol. 2011. PMID: 21969385 Free PMC article.
-
Genomics and transcriptomics to protect rice (Oryza sativa. L.) from abiotic stressors: -pathways to achieving zero hunger.Front Plant Sci. 2022 Oct 20;13:1002596. doi: 10.3389/fpls.2022.1002596. eCollection 2022. Front Plant Sci. 2022. PMID: 36340401 Free PMC article. Review.
-
Abiotic Stress in Rice: Visiting the Physiological Response and Its Tolerance Mechanisms.Plants (Basel). 2023 Nov 23;12(23):3948. doi: 10.3390/plants12233948. Plants (Basel). 2023. PMID: 38068585 Free PMC article. Review.
Cited by
-
Post-Translational Modifications in Histones and Their Role in Abiotic Stress Tolerance in Plants.Proteomes. 2023 Nov 22;11(4):38. doi: 10.3390/proteomes11040038. Proteomes. 2023. PMID: 38133152 Free PMC article. Review.
-
OsPUB41, a U-box E3 ubiquitin ligase, acts as a negative regulator of drought stress response in rice (Oryza Sativa L.).Plant Mol Biol. 2021 Jul;106(4-5):463-477. doi: 10.1007/s11103-021-01158-4. Epub 2021 Jun 7. Plant Mol Biol. 2021. PMID: 34100185
-
E3 ligase, the Oryza sativa salt-induced RING finger protein 4 (OsSIRP4), negatively regulates salt stress responses via degradation of the OsPEX11-1 protein.Plant Mol Biol. 2021 Feb;105(3):231-245. doi: 10.1007/s11103-020-01084-x. Epub 2020 Oct 20. Plant Mol Biol. 2021. PMID: 33079323
-
Deciphering the mechanism of E3 ubiquitin ligases in plant responses to abiotic and biotic stresses and perspectives on PROTACs for crop resistance.Plant Biotechnol J. 2024 Oct;22(10):2811-2843. doi: 10.1111/pbi.14407. Epub 2024 Jun 12. Plant Biotechnol J. 2024. PMID: 38864414 Free PMC article. Review.
-
Proteomes and ubiquitylomes reveal the regulation mechanism of cold tolerance mediated by OsGRF4 in rice.Front Plant Sci. 2025 Mar 21;16:1531399. doi: 10.3389/fpls.2025.1531399. eCollection 2025. Front Plant Sci. 2025. PMID: 40190655 Free PMC article.
References
-
- Bueso E., Rodriguez L., Lorenzo-Orts L., Gonzalez-Guzman M., Sayas E., Muñoz-Bertomeu J., et al. (2014). The single-subunit RING-type E3 ubiquitin ligase RSL1 targets PYL4 and PYR1 ABA receptors in plasma membrane to modulate abscisic acid signaling. Plant J. 80 1057–1071. 10.1111/tpj.12708 - DOI - PubMed
-
- Byun M. Y., Cui L. H., Lee J., Park H., Lee A., Kim W. T., et al. (2018). Identification of rice genes associated with enhanced cold tolerance by comparative transcriptome analysis with two transgenic rice plants overexpressing DaCBF4 and DaCBF7 isolated from Antarctic flowering plant Deschamsia antarctica. Front. Plant Sci. 9:601. 10.3389/fpls.2018.00601 - DOI - PMC - PubMed
-
- Byun M. Y., Cui L. H., Oh T. K., Jung Y., Lee A., Park K. Y., et al. (2017). Homologous U-box E3 ubiquitin ligases OsPUB2 and OsPUB3 are involved in the positive regulation of low temperature stress response in rice (Oryza sativa L.). Front. Plant Sci. 8:16. 10.3389/fpls.2017.00016 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Research Materials