

Autophagy: An Essential Degradation Program for Cellular Homeostasis and Life
- PMID: 30572663
- PMCID: PMC6315530
- DOI: 10.3390/cells7120278
Autophagy: An Essential Degradation Program for Cellular Homeostasis and Life
Abstract
Autophagy is a lysosome-dependent cellular degradation program that responds to a variety of environmental and cellular stresses. It is an evolutionarily well-conserved and essential pathway to maintain cellular homeostasis, therefore, dysfunction of autophagy is closely associated with a wide spectrum of human pathophysiological conditions including cancers and neurodegenerative diseases. The discovery and characterization of the kingdom of autophagy proteins have uncovered the molecular basis of the autophagy process. In addition, recent advances on the various post-translational modifications of autophagy proteins have shed light on the multiple layers of autophagy regulatory mechanisms, and provide novel therapeutic targets for the treatment of the diseases.
Keywords: AMPK-mTORC1-ULK1 triad; PIK3C3/VPS34; autophagy; autophagy adaptor; regulatory modifications.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
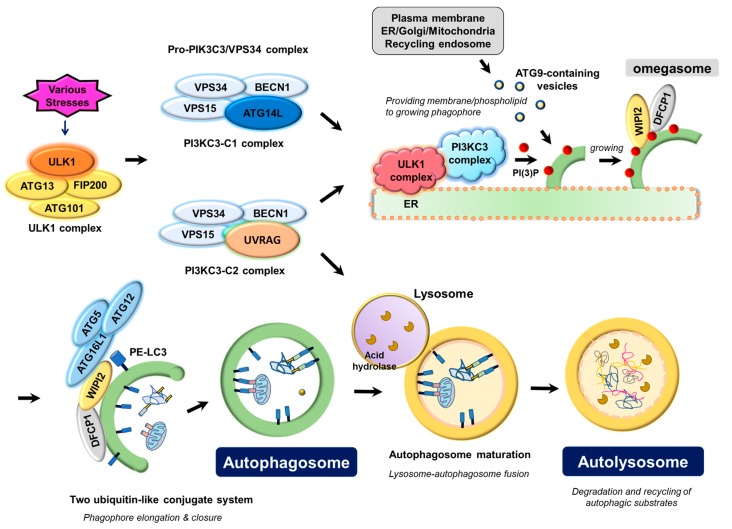
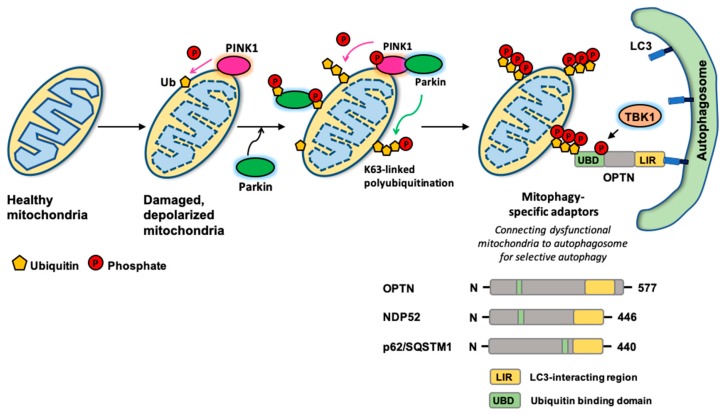
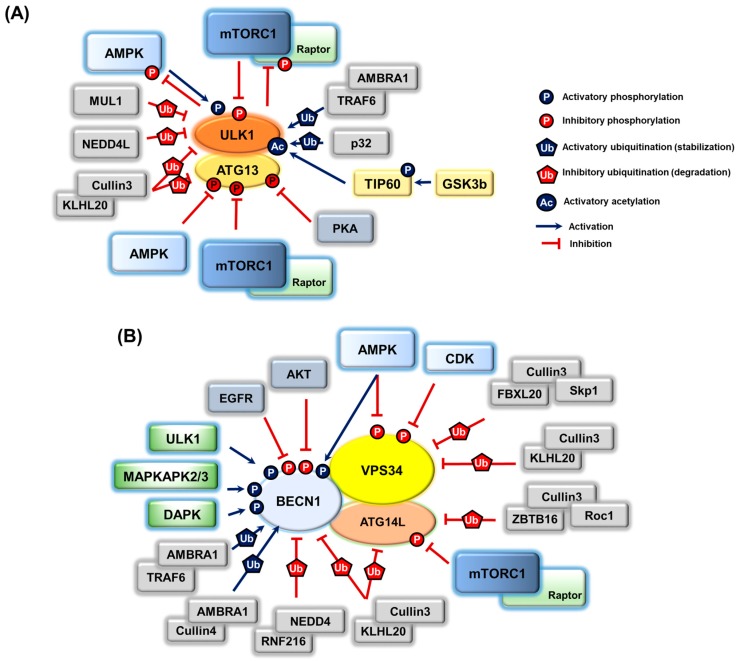
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
