

Flavin-based metabolic cycles are integral features of growth and division in single yeast cells
- PMID: 30575765
- PMCID: PMC6303410
- DOI: 10.1038/s41598-018-35936-w
Flavin-based metabolic cycles are integral features of growth and division in single yeast cells
Abstract
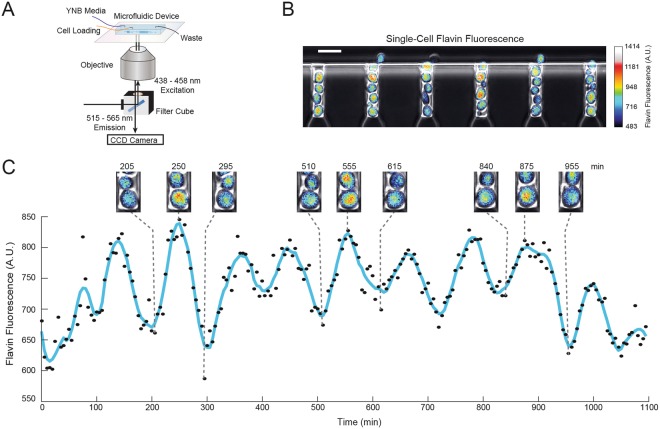
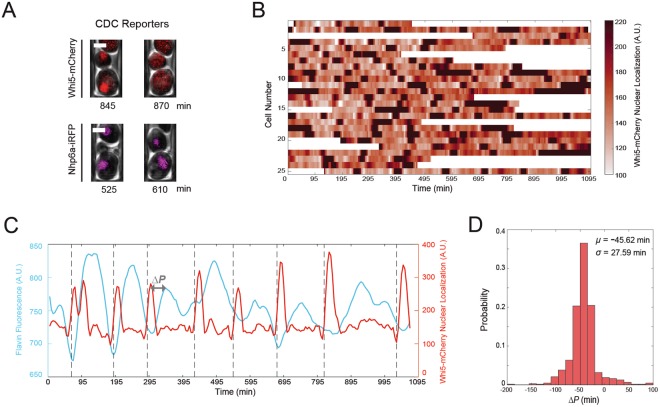
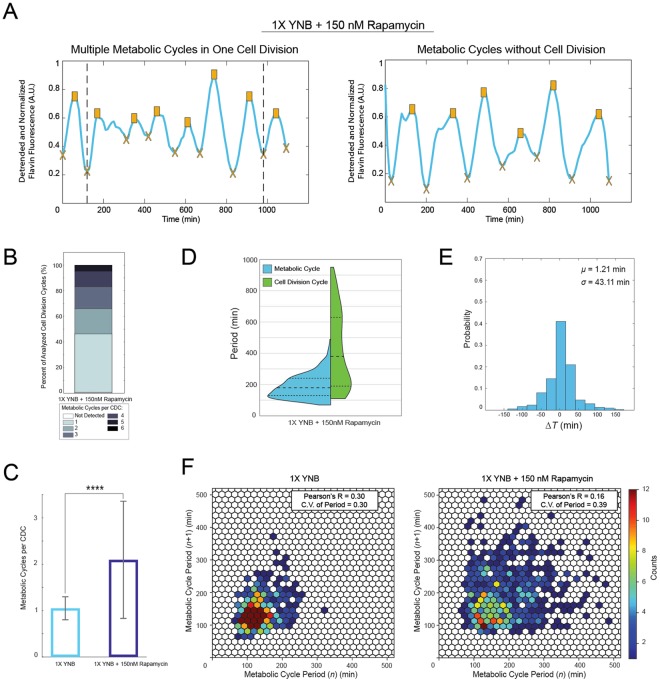
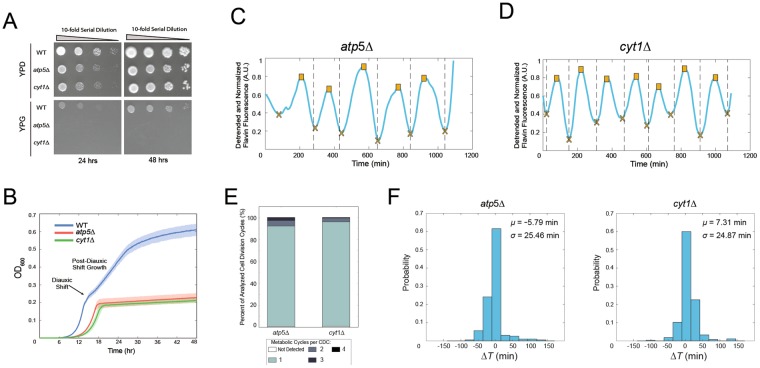
The yeast metabolic cycle (YMC) is a fascinating example of biological organization, in which cells constrain the function of specific genetic, protein and metabolic networks to precise temporal windows as they grow and divide. However, understanding the intracellular origins of the YMC remains a challenging goal, as measuring the oxygen oscillations traditionally associated with it requires the use of synchronized cultures growing in nutrient-limited chemostat environments. To address these limitations, we used custom-built microfluidic devices and time-lapse fluorescence microscopy to search for metabolic cycling in the form of endogenous flavin fluorescence in unsynchronized single yeast cells. We uncovered robust and pervasive metabolic cycles that were synchronized with the cell division cycle (CDC) and oscillated across four different nutrient conditions. We then studied the response of these metabolic cycles to chemical and genetic perturbations, showing that their phase synchronization with the CDC can be altered through treatment with rapamycin, and that metabolic cycles continue even in respiratory deficient strains. These results provide a foundation for future studies of the physiological importance of metabolic cycles in processes such as CDC control, metabolic regulation and cell aging.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Mirollo RE, Strogatz SH. Synchronization of pulse-coupled biological oscillators. SIAM J. on Appl. Math. 1990;50:1645–1662. doi: 10.1137/0150098. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
