

Toll-Like Receptor-4 Disruption Suppresses Adipose Tissue Remodeling and Increases Survival in Cancer Cachexia Syndrome
- PMID: 30575787
- PMCID: PMC6303407
- DOI: 10.1038/s41598-018-36626-3
Toll-Like Receptor-4 Disruption Suppresses Adipose Tissue Remodeling and Increases Survival in Cancer Cachexia Syndrome
Abstract
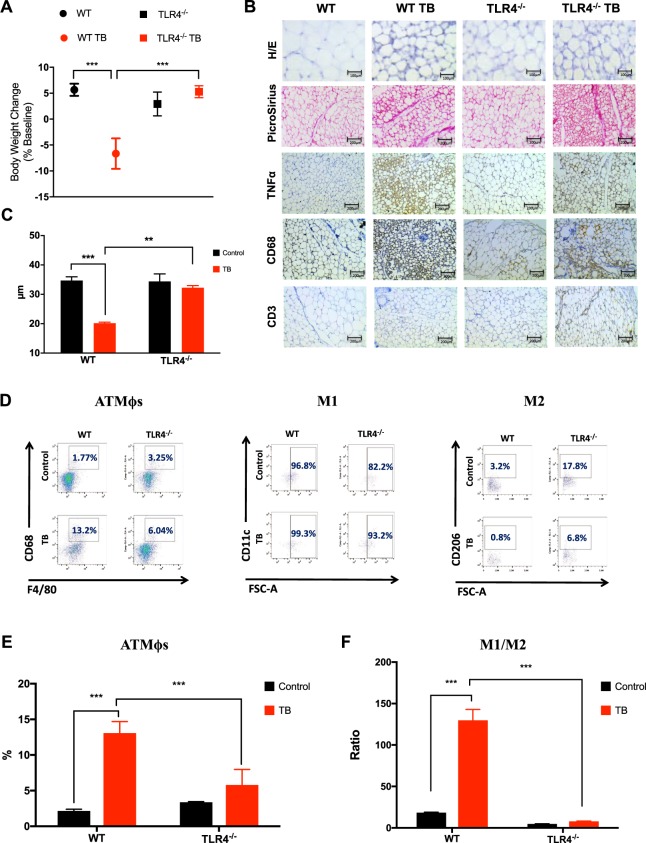
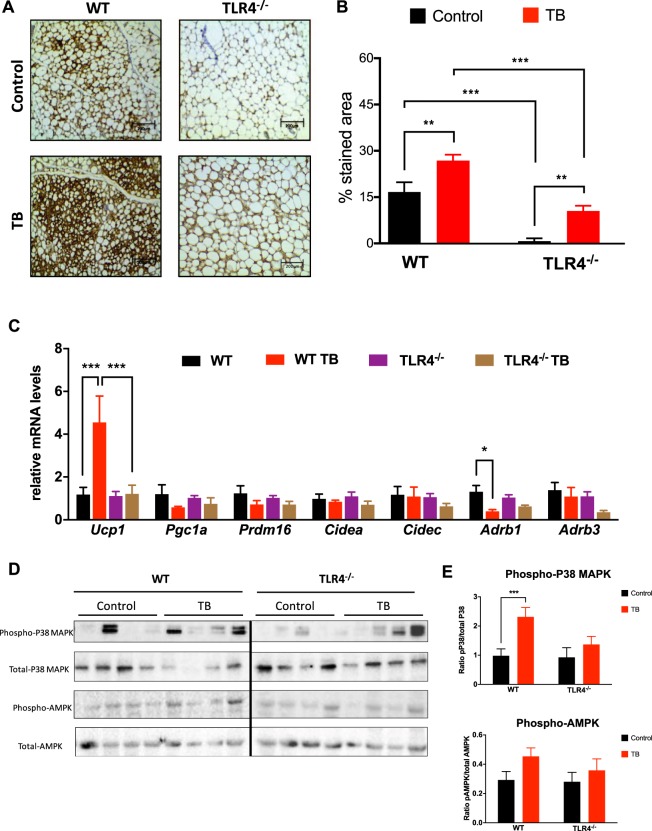
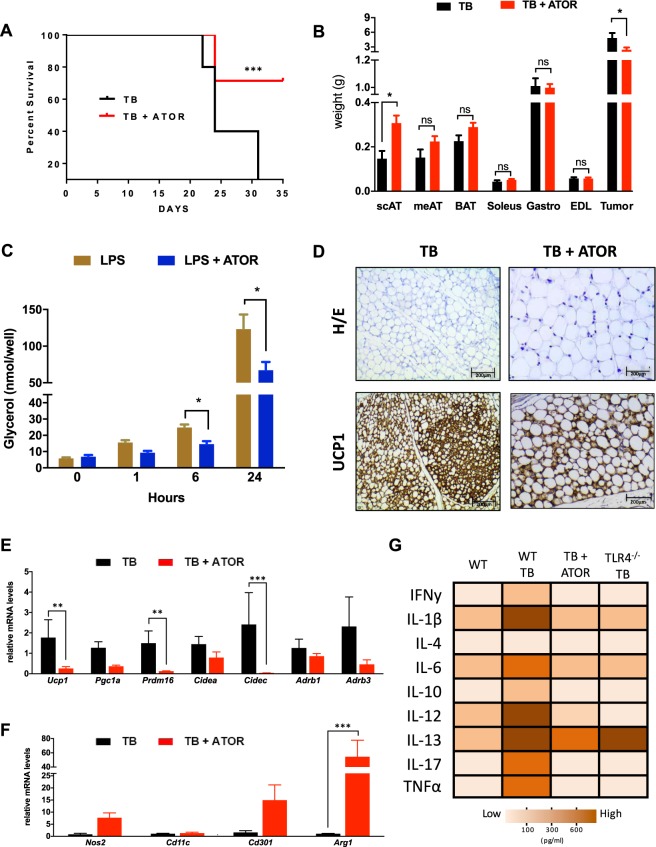
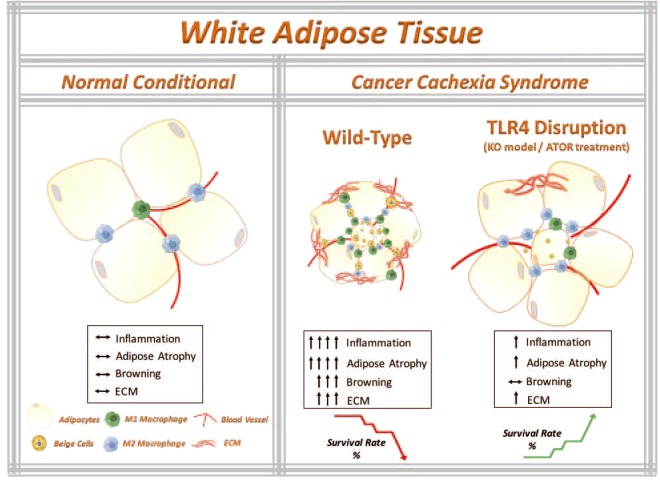
Cancer-induced cachexia, characterized by systemic inflammation, body weight loss, adipose tissue (AT) remodeling and muscle wasting, is a malignant metabolic syndrome with undefined etiology. Here, we show that both genetic ablation and pharmacological inhibition of TLR4 were able to attenuate the main clinical markers of cachexia in mice bearing Lewis lung carcinoma (LLC). AT remodelling was not found in LLC tumor-bearing (TB) TLR4-/- mice due to reduced macrophage infiltration and adipocyte atrophy. TLR4-/- mice were also resistant to cold-induced browning of subcutaneous AT (scAT). Importantly, pharmacological inhibition of TLR4 (Atorvastatin) reproduced the main protective effect against AT remodeling found in TLR4-/- TB mice. Moreover, the treatment was effective in prolonging survival and attenuating tumor mass growth when compared to non-treated-TB animals. Furthermore, tumor-induced elevation of circulating pro-inflammatory cytokines was similarly abolished in both genetic ablation and pharmacological inhibition of TLR4. These data suggest that TLR4 is a critical mediator and a promising target for novel anti-cachexia therapies.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
