

Effects of RNA structure and salt concentration on the affinity and kinetics of interactions between pentatricopeptide repeat proteins and their RNA ligands
- PMID: 30576379
- PMCID: PMC6303017
- DOI: 10.1371/journal.pone.0209713
Effects of RNA structure and salt concentration on the affinity and kinetics of interactions between pentatricopeptide repeat proteins and their RNA ligands
Abstract
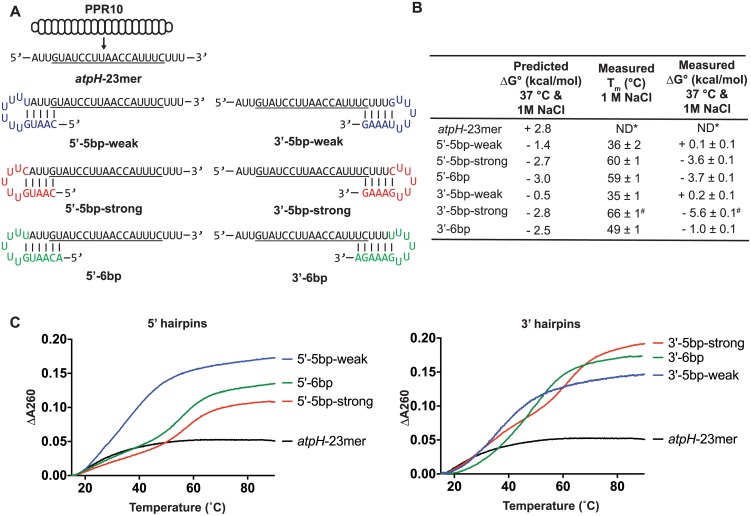
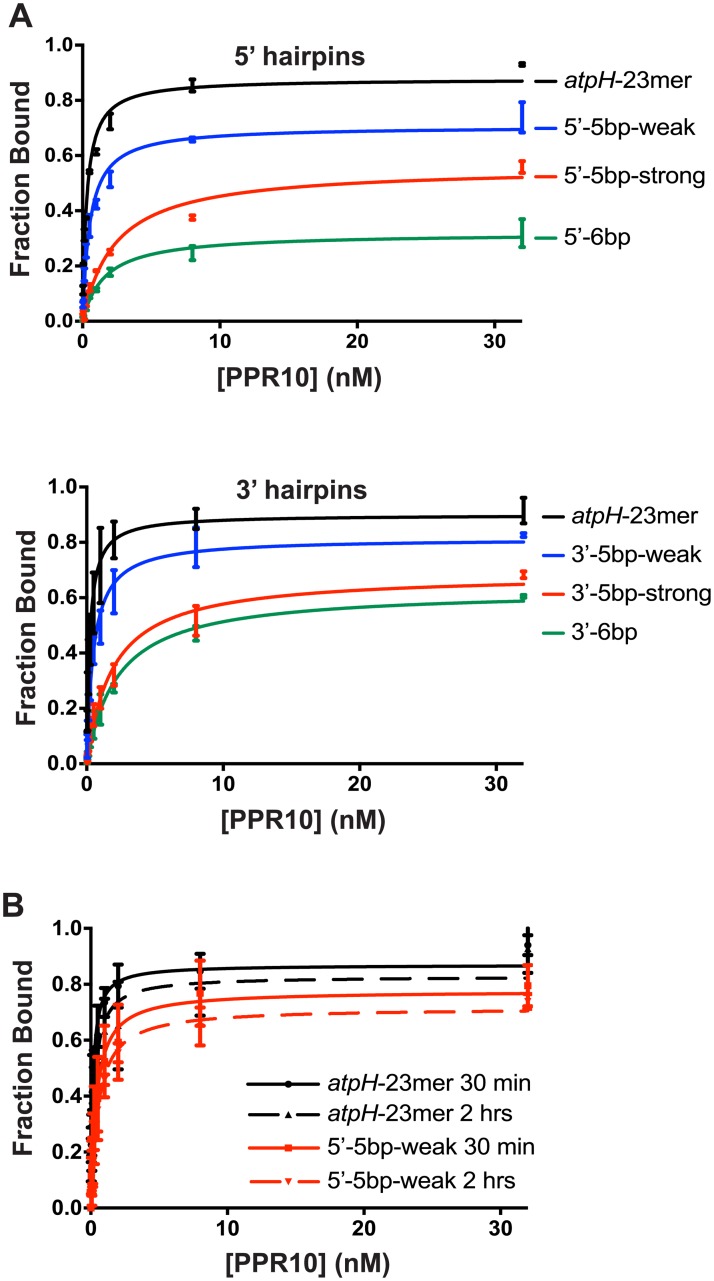
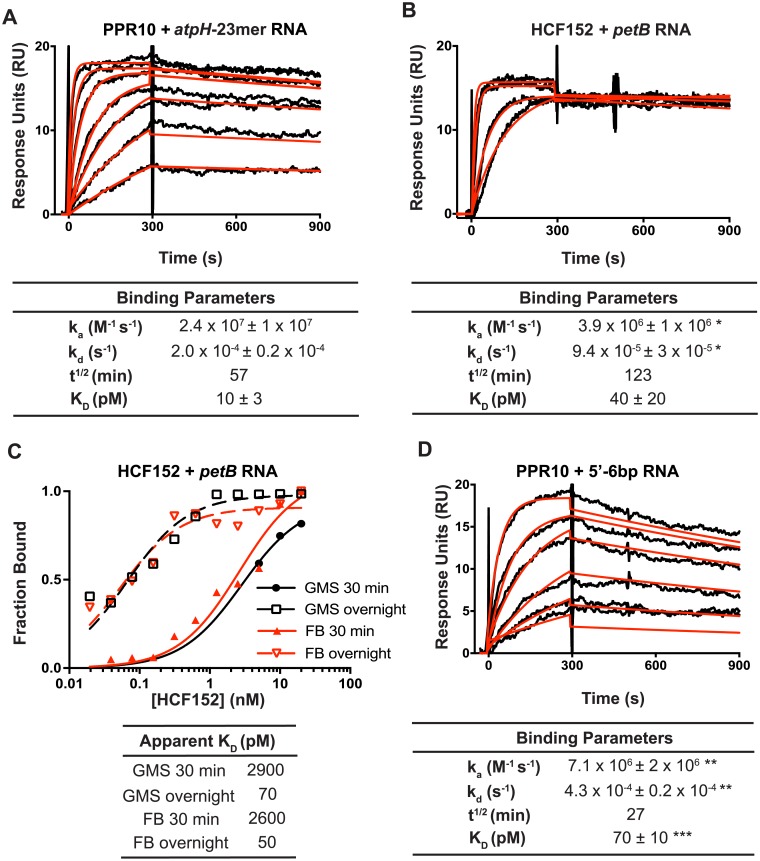
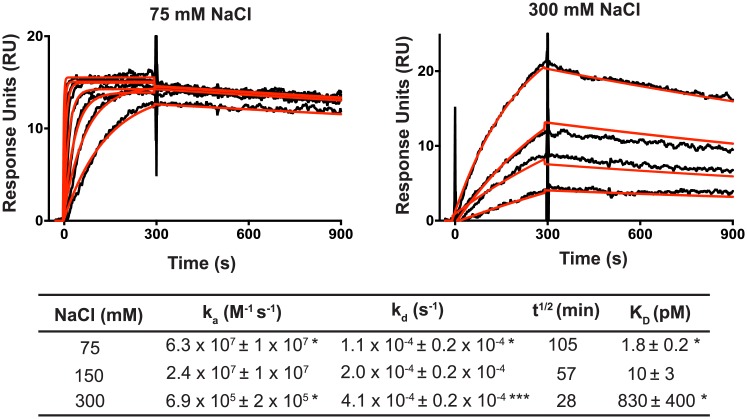
Pentatricopeptide repeat (PPR) proteins are helical repeat proteins that bind specific RNA sequences via modular 1-repeat:1-nucleotide interactions. Binding specificity is dictated, in part, by hydrogen bonds between the amino acids at two positions in each PPR motif and the Watson-Crick face of the aligned nucleobase. There is evidence that PPR-RNA interactions can compete with RNA-RNA interactions in vivo, and that this competition underlies some effects of PPR proteins on gene expression. Conversely, RNA secondary structure can inhibit the binding of a PPR protein to its specific binding site. The parameters that influence whether PPR-RNA or RNA-RNA interactions prevail are unknown. Understanding these parameters will be important for understanding the functions of natural PPR proteins and for the design of engineered PPR proteins for synthetic biology purposes. We addressed this question by analyzing the effects of RNA structures of varying stability and position on the binding of the model protein PPR10 to its atpH RNA ligand. Our results show that even very weak RNA structures (ΔG° ~ 0 kcal/mol) involving only one nucleotide at either end of the minimal binding site impede PPR10 binding. Analysis of binding kinetics using Surface Plasmon Resonance showed that RNA structures reduce PPR10's on-rate and increase its off-rate. Complexes between the PPR proteins PPR10 and HCF152 and their respective RNA ligands have long half-lives (one hour or more), correlating with their functions as barriers to exonucleolytic RNA decay in vivo. The effects of salt concentration on PPR10-RNA binding kinetics showed that electrostatic interactions play an important role in establishing PPR10-RNA interactions but play a relatively small role in maintaining specific interactions once established.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Small I, Peeters N. The PPR motif—a TPR-related motif prevalent in plant organellar proteins. Trends Biochem Sci 2000; 25: 46–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
