

Genome-Wide Analysis, Expression Profile, and Characterization of the Acid Invertase Gene Family in Pepper
- PMID: 30577540
- PMCID: PMC6337152
- DOI: 10.3390/ijms20010015
Genome-Wide Analysis, Expression Profile, and Characterization of the Acid Invertase Gene Family in Pepper
Abstract
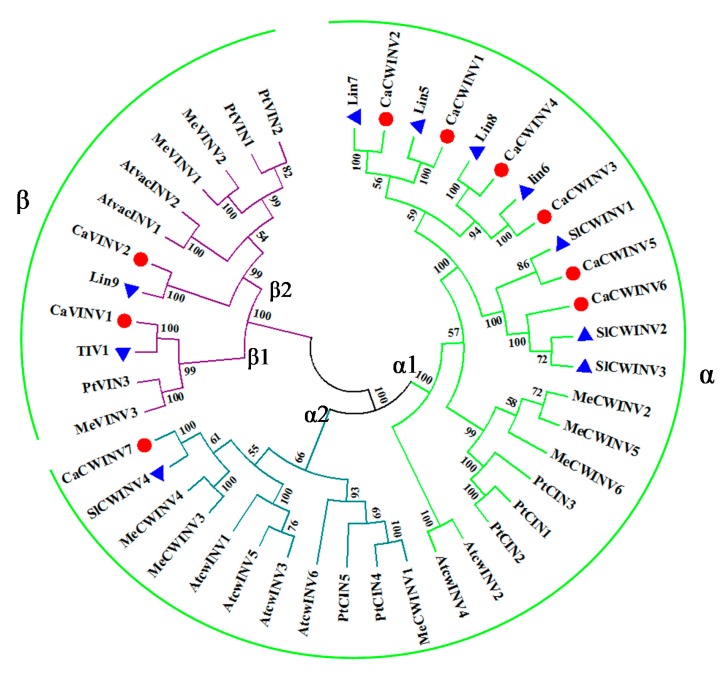
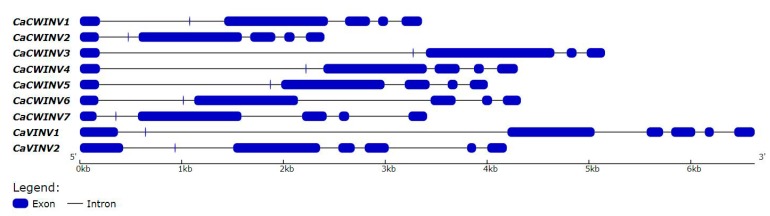
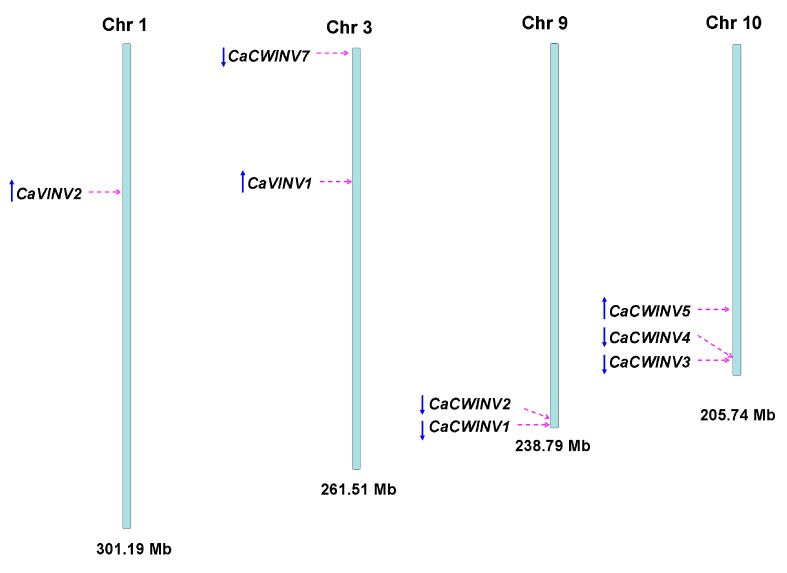
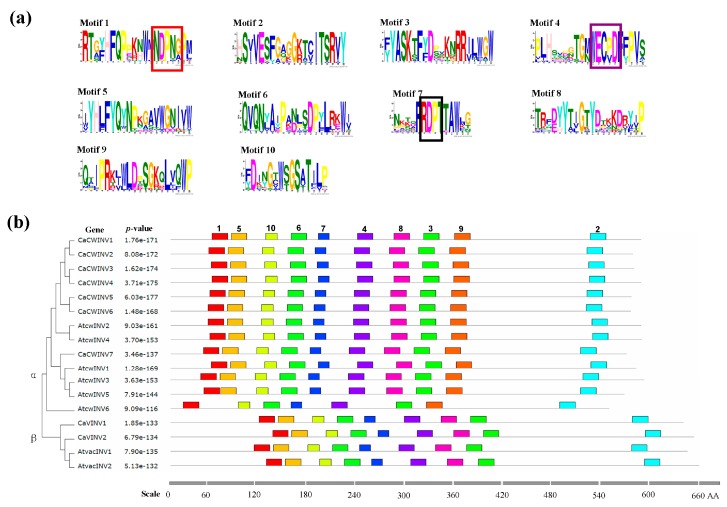
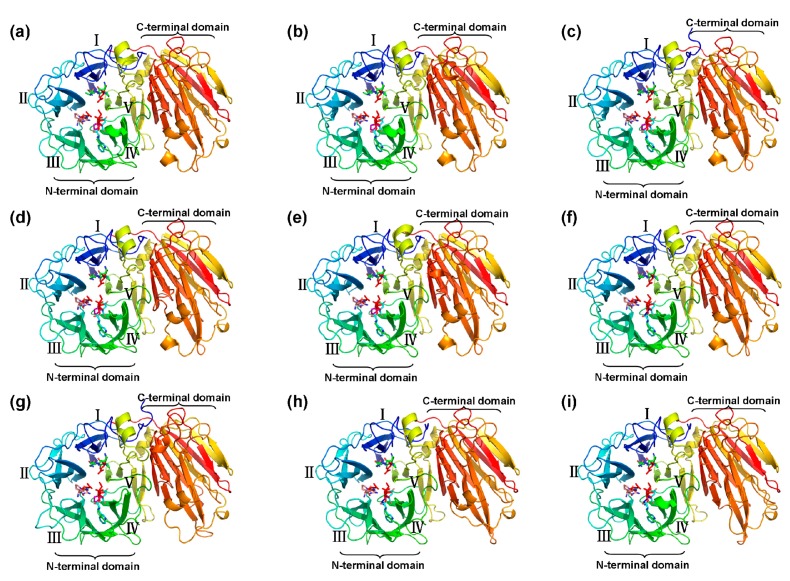
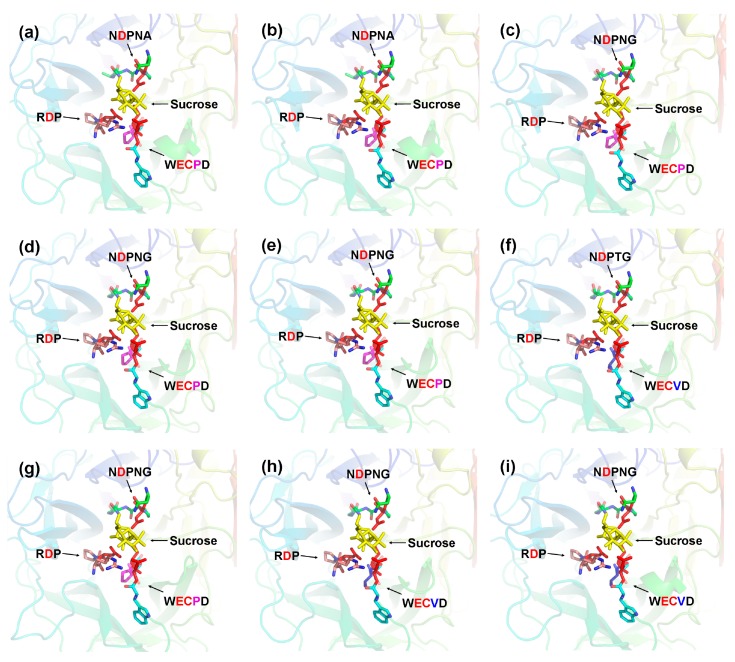
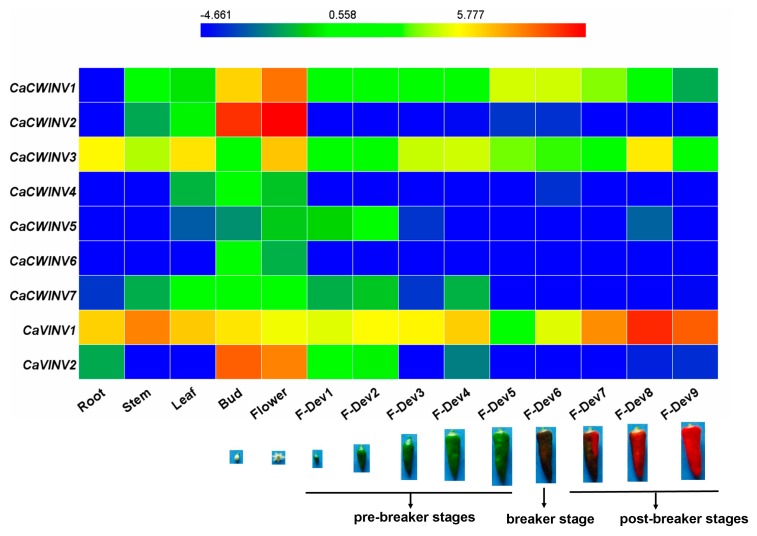
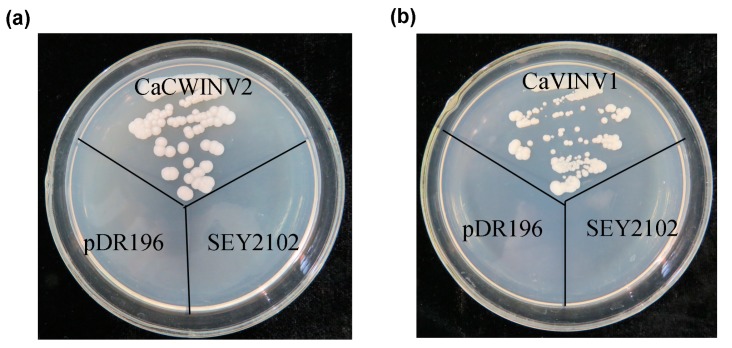
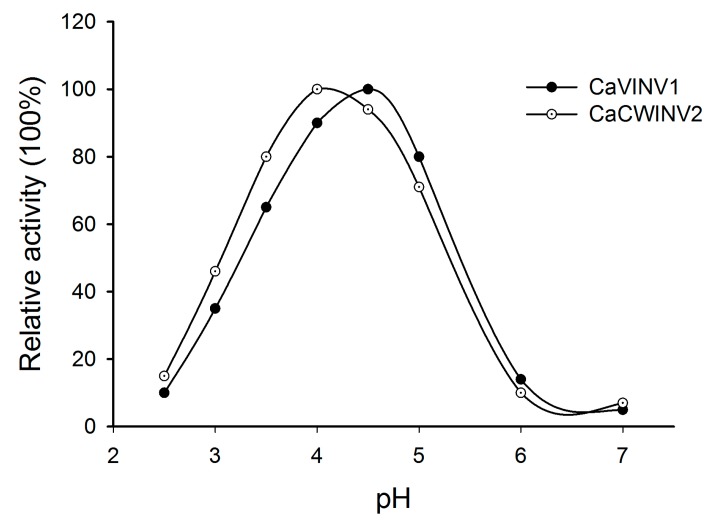
Catalytic decomposition of sucrose by acid invertases (AINVs) under acidic conditions plays an important role in the development of sink organs in plants. To reveal the function of AINVs in the development of pepper fruits, nine AINV genes of pepper were identified. Protein sequencing and phylogenetic analysis revealed that the CaAINV family may be divided into cell wall invertases (CaCWINV1⁻7) and vacuolar invertases (CaVINV1⁻2). CaAINVs contain conserved regions and protein structures typical of the AINVs in other plants. Gene expression profiling indicated that CaCWINV2 and CaVINV1 were highly expressed in reproductive organs but differed in expression pattern. CaCWINV2 was mainly expressed in buds and flowers, while CaVINV1 was expressed in developmental stages, such as the post-breaker stage. Furthermore, invertase activity of CaCWINV2 and CaVINV1 was identified via functional complementation in an invertase-deficient yeast. Optimum pH for CaCWINV2 and CaVINV1 was found to be 4.0 and 4.5, respectively. Gene expression and enzymatic activity of CaCWINV2 and CaVINV1 indicate that these AINV enzymes may be pivotal for sucrose hydrolysis in the reproductive organs of pepper.
Keywords: acid invertase; enzymatic activity; gene expression; pepper.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Elwan M.W.M., El-Hamahmy M.A.M. Improved productivity and quality associated with salicylic acid application in greenhouse pepper. Sci. Hortic. 2009;122:521–526. doi: 10.1016/j.scienta.2009.07.001. - DOI
-
- Aloni B., Karni L., Zaidman Z., Schaffer A.A. Changes of Carbohydrates in Pepper (Capsicum annuum L.) Flowers in Relation to Their Abscission Under Different Shading Regimes. Ann. Bot. 1996;78:163–168. doi: 10.1006/anbo.1996.0109. - DOI
-
- Hubbard N.L., Pharr D.M. Developmental changes in carbohydrate concentration and activaties of sucrose metabolizing enzymes in fruits of two Capsicum annuum L. genotypes. Plant Sci. 1992;86:33–39. doi: 10.1016/0168-9452(92)90176-M. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
