

Cell-type and projection-specific dopaminergic encoding of aversive stimuli in addiction
- PMID: 30580012
- PMCID: PMC6506354
- DOI: 10.1016/j.brainres.2018.12.024
Cell-type and projection-specific dopaminergic encoding of aversive stimuli in addiction
Abstract
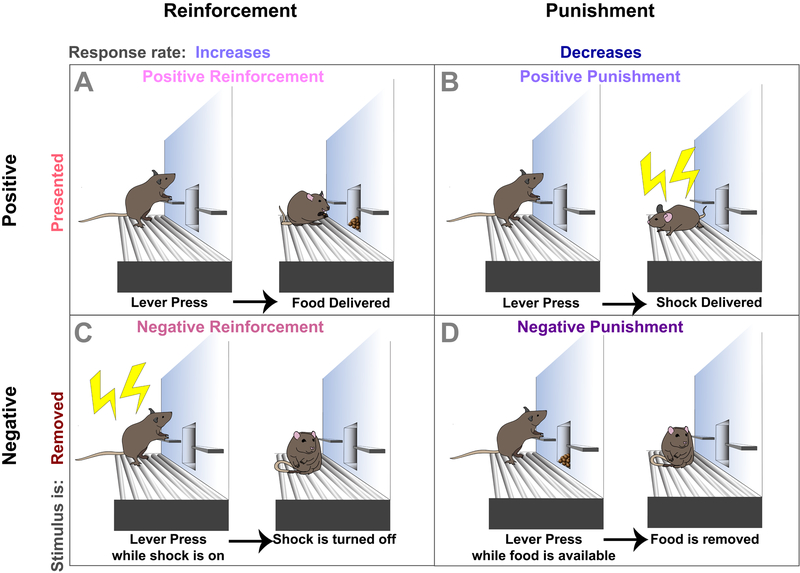
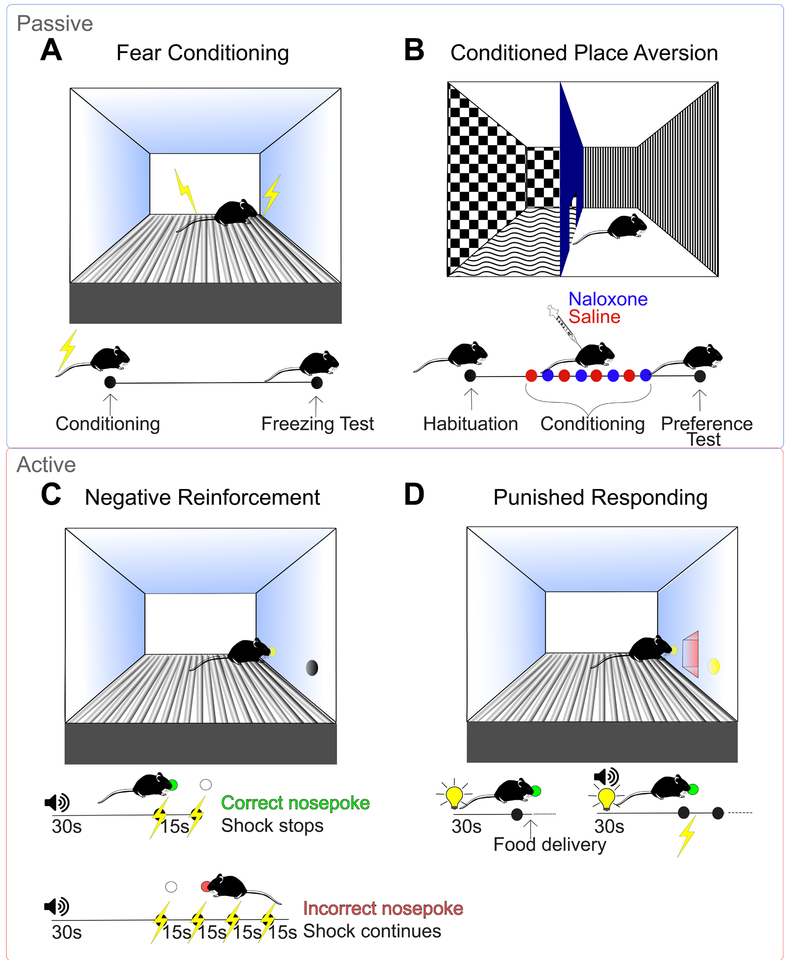
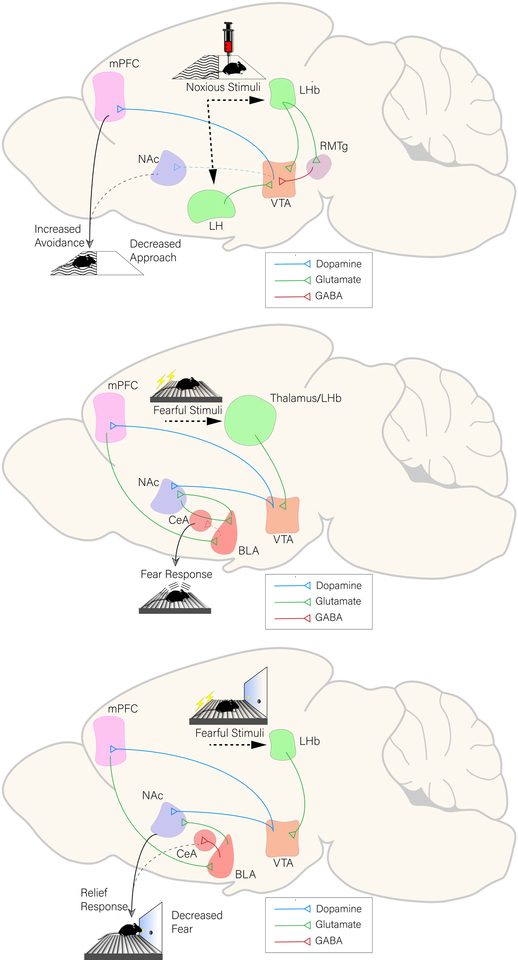
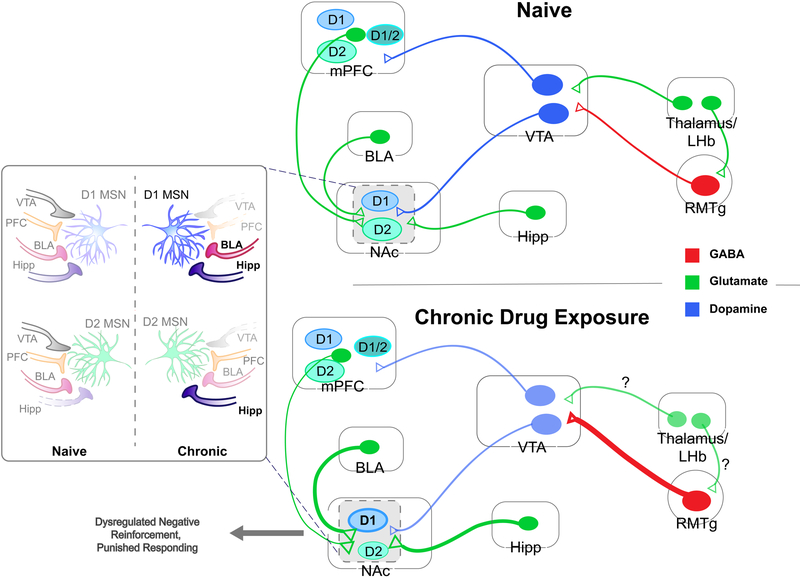
Drug addiction is a major public health concern across the world for which there are limited treatment options. In order to develop new therapies to correct the behavioral deficits that result from repeated drug use, we need to understand the neural circuit dysfunction that underlies the pathophysiology of the disorder. Because the initial reinforcing effects of drugs are dependent on increases in dopamine in reward-related brain regions such as the mesolimbic dopamine pathway, a large focus of addiction research has centered on the dysregulation of this system and its control of positive reinforcement and motivation. However, in addition to the processing of positive, rewarding stimuli, there are clear deficits in the encoding and valuation of information about potential negative outcomes and how they control decision making and motivation. Further, aversive stimuli can motivate or suppress behavior depending on the context in which they are encountered. We propose a model where rewarding and aversive information guides the execution of specific motivated actions through mesocortical and mesolimbic dopamine acting on D1- and D2- receptor containing neuronal populations. Volitional drug exposure alters the processing of rewarding and aversive stimuli through remodeling of these dopaminergic circuits, causing maladaptive drug seeking, self-administration in the face of negative consequences, and drug craving. Together, this review discusses the dysfunction of the circuits controlling different types of aversive learning as well as how these guide specific discrete behaviors, and provides a conceptual framework for how they should be considered in preclinical addiction models.
Keywords: Associative learning; Cocaine; Nucleus accumbens; Prefrontal cortex; Punishment; Self-administration.
Copyright © 2018 Elsevier B.V. All rights reserved.
Figures
Similar articles
-
Neural substrates of drug craving and relapse in drug addiction.Ann Med. 1998 Aug;30(4):379-89. doi: 10.3109/07853899809029938. Ann Med. 1998. PMID: 9783837 Review.
-
Addiction and brain reward and antireward pathways.Adv Psychosom Med. 2011;30:22-60. doi: 10.1159/000324065. Epub 2011 Apr 19. Adv Psychosom Med. 2011. PMID: 21508625 Free PMC article. Review.
-
Drug addiction. Part II. Neurobiology of addiction.Pol J Pharmacol. 2001 Jul-Aug;53(4):303-17. Pol J Pharmacol. 2001. PMID: 11990077 Review.
-
Neurobiologic processes in drug reward and addiction.Harv Rev Psychiatry. 2004 Nov-Dec;12(6):305-20. doi: 10.1080/10673220490910844. Harv Rev Psychiatry. 2004. PMID: 15764467 Free PMC article. Review.
-
Neural mechanisms of the nucleus accumbens circuit in reward and aversive learning.Neurosci Res. 2016 Jul;108:1-5. doi: 10.1016/j.neures.2016.01.004. Epub 2016 Jan 28. Neurosci Res. 2016. PMID: 26827817 Review.
Cited by
-
Distinct Role of Dopamine in the PFC and NAc During Exposure to Cocaine-Associated Cues.Int J Neuropsychopharmacol. 2021 Dec 8;24(12):988-1001. doi: 10.1093/ijnp/pyab067. Int J Neuropsychopharmacol. 2021. PMID: 34626116 Free PMC article.
-
Sex differences in dopamine release regulation in the striatum.Neuropsychopharmacology. 2021 Feb;46(3):491-499. doi: 10.1038/s41386-020-00915-1. Epub 2020 Dec 14. Neuropsychopharmacology. 2021. PMID: 33318634 Free PMC article. Review.
-
Lateral Orbitofrontal Cortex Encodes Presence of Risk and Subjective Risk Preference During Decision-Making.bioRxiv [Preprint]. 2024 Apr 9:2024.04.08.588332. doi: 10.1101/2024.04.08.588332. bioRxiv. 2024. Update in: Cereb Cortex. 2025 Jun 4;35(6):bhaf146. doi: 10.1093/cercor/bhaf146. PMID: 38645204 Free PMC article. Updated. Preprint.
-
Midbrain circuits of novelty processing.Neurobiol Learn Mem. 2020 Dec;176:107323. doi: 10.1016/j.nlm.2020.107323. Epub 2020 Oct 11. Neurobiol Learn Mem. 2020. PMID: 33053429 Free PMC article. Review.
-
D1 and D2 medium spiny neurons in the nucleus accumbens core have distinct and valence-independent roles in learning.Neuron. 2024 Mar 6;112(5):835-849.e7. doi: 10.1016/j.neuron.2023.11.023. Epub 2023 Dec 21. Neuron. 2024. PMID: 38134921 Free PMC article.
References
-
- Abercrombie ED, Keefe KA, DiFrischia DS, & Zigmond MJ (1989). Differential effect of stress on in vivo dopamine release in striatum, nucleus accumbens, and medial frontal cortex. Journal of Neurochemistry, 52, 1655–1658. - PubMed
-
- Al-Hasani R, McCall JG, Shin G, Gomez AM, Schmitz GP, Bernardi JM, Pyo C-O, Park SI, Marcinkiewcz CM, Crowley NA, Krashes MJ, Lowell BB, Kash TL, Rogers JA, Bruchas MR, 2015. Distinct Subpopulations of Nucleus Accumbens Dynorphin Neurons Drive Aversion and Reward. Neuron 87, 1063–1077. - PMC - PubMed
