

Repair of protein-linked DNA double strand breaks: Using the adenovirus genome as a model substrate in cell-based assays
- PMID: 30583959
- PMCID: PMC8057058
- DOI: 10.1016/j.dnarep.2018.12.001
Repair of protein-linked DNA double strand breaks: Using the adenovirus genome as a model substrate in cell-based assays
Abstract
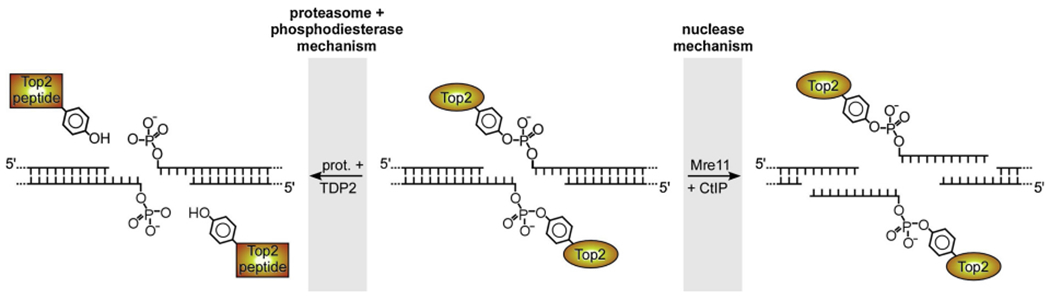
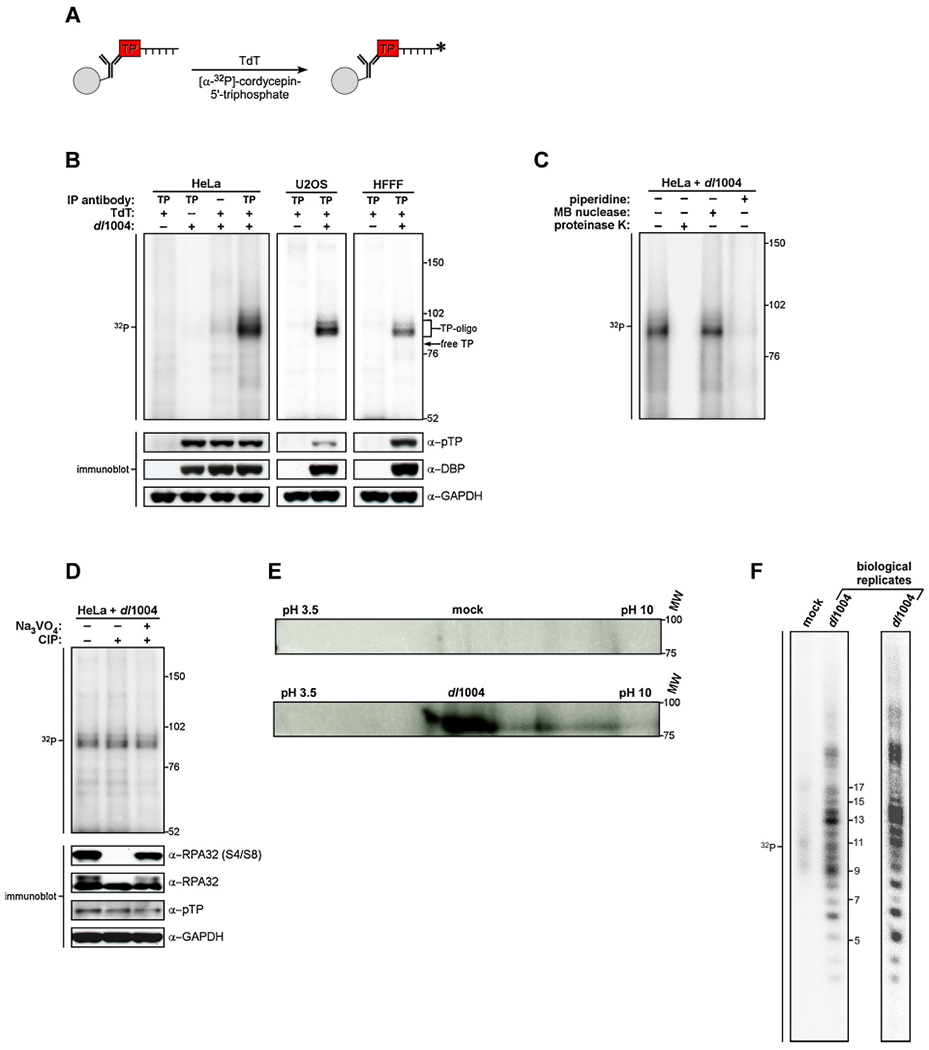
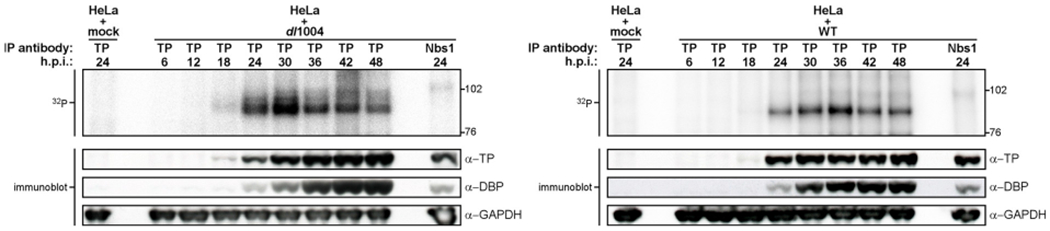
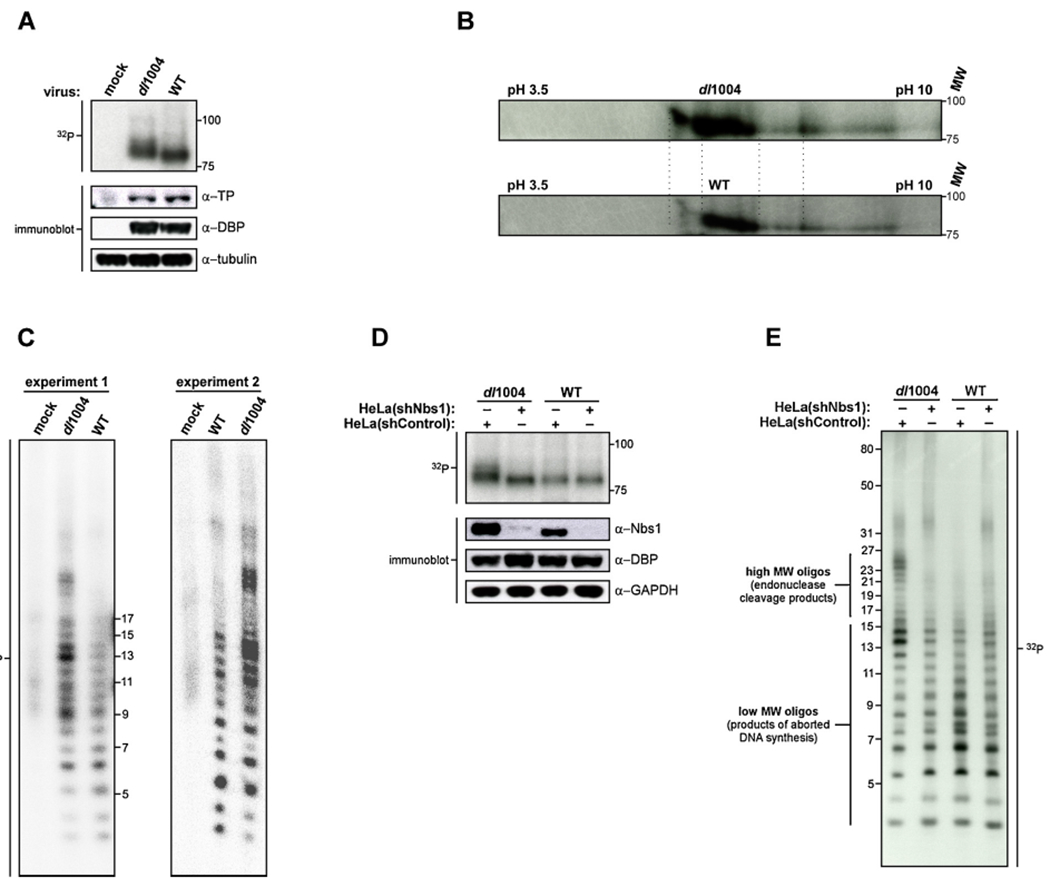
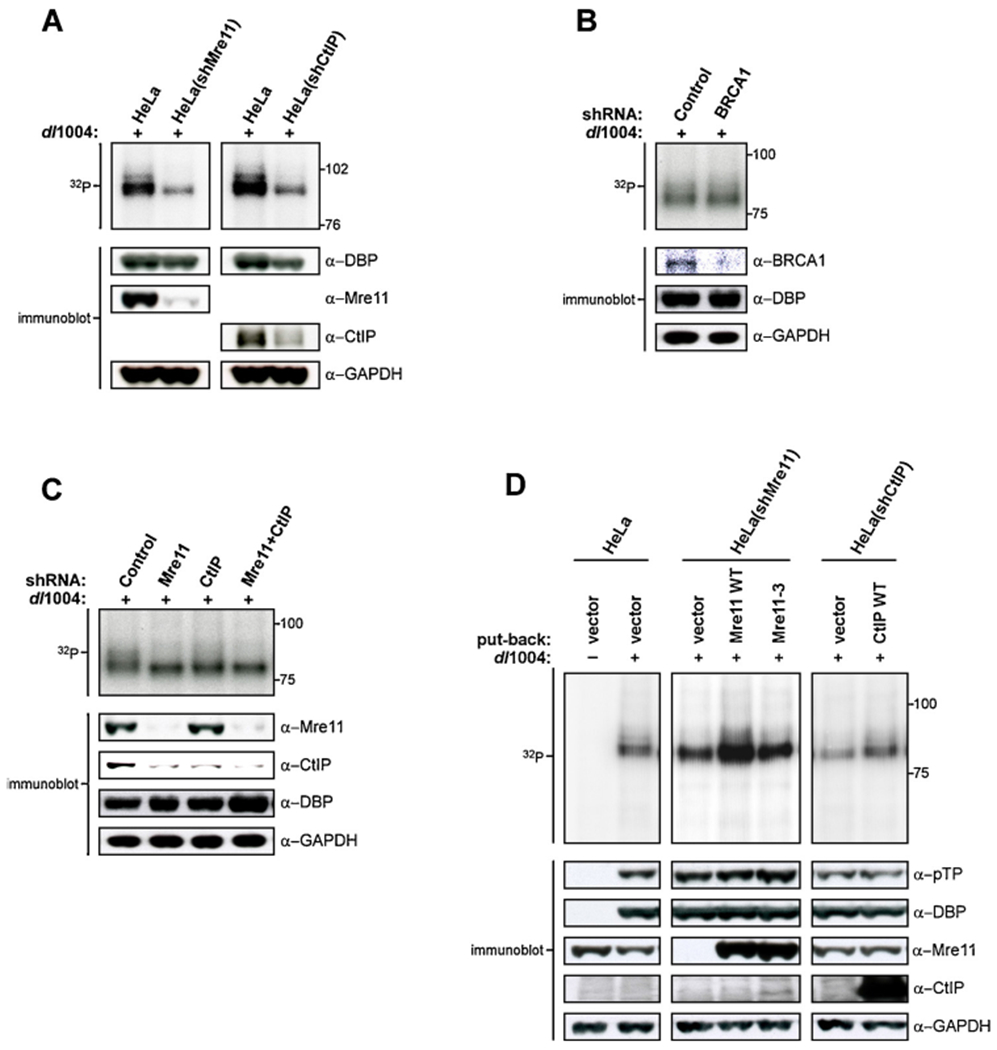
The DNA double strand breaks (DSBs) created during meiotic recombination and during some types of chemotherapy contain protein covalently attached to their 5' termini. Removal of the end-blocking protein is a prerequisite to DSB processing by non-homologous end-joining or homologous recombination. One mechanism for removing the protein involves CtIP-stimulated Mre11-catalyzed nicking of the protein-linked strand distal to the DSB terminus, releasing the end-blocking protein while it remains covalently attached to an oligonucleotide. Much of what is known about this repair process has recently been deciphered through in vitro reconstitution studies. We present here a novel model system based on adenovirus (Ad), which contains the Ad terminal protein covalently linked to the 5' terminus of its dsDNA genome, for studying the repair of 5' protein-linked DSBs in vivo. It was previously shown that the genome of Ad mutants that lack early region 4 (E4) can be joined into concatemers in vivo, suggesting that the Ad terminal protein had been removed from the genome termini prior to ligation. Here we show that during infection with the E4-deleted Ad mutant dl1004, the Ad terminal protein is removed in a manner that recapitulates removal of end-blocking proteins from cellular DSBs. In addition to displaying a dependence on CtIP, and Mre11 acting as the endonuclease, the protein-linked oligonucleotides that are released from the viral genome are similar in size to the oligos that remain attached to Spo11 and Top2 after they are removed from the 5' termini of DSBs during meiotic recombination and etoposide chemotherapy, respectively. The single nucleotide resolution that is possible with this assay, combined with the single sequence context in which the lesion is presented, make it a useful tool for further refining our mechanistic understanding of how blocking proteins are removed from the 5' termini of DSBs.
Keywords: Adenovirus; Chemotherapy; Concatemer; CtIP; DNA double strand break; End blocking; End processing; Endonuclease; Etoposide; Homologous recombination; Ligation; MRN; Mre11; Nonhomologous end joining; Protein-DNA adduct; Protein-linked DSB; Spo11; Terminal protein; Topoisomerase 2; dl1004.
Copyright © 2018. Published by Elsevier B.V.
Conflict of interest statement
Conflicts of interest
The authors declare that this research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
CtIP and MRN promote non-homologous end-joining of etoposide-induced DNA double-strand breaks in G1.Nucleic Acids Res. 2011 Mar;39(6):2144-52. doi: 10.1093/nar/gkq1175. Epub 2010 Nov 17. Nucleic Acids Res. 2011. PMID: 21087997 Free PMC article.
-
CtIP-BRCA1 modulates the choice of DNA double-strand-break repair pathway throughout the cell cycle.Nature. 2009 May 21;459(7245):460-3. doi: 10.1038/nature07955. Epub 2009 Apr 8. Nature. 2009. PMID: 19357644 Free PMC article.
-
Cell cycle-dependent complex formation of BRCA1.CtIP.MRN is important for DNA double-strand break repair.J Biol Chem. 2008 Mar 21;283(12):7713-20. doi: 10.1074/jbc.M710245200. Epub 2008 Jan 2. J Biol Chem. 2008. PMID: 18171670
-
Type II DNA Topoisomerases Cause Spontaneous Double-Strand Breaks in Genomic DNA.Genes (Basel). 2019 Oct 30;10(11):868. doi: 10.3390/genes10110868. Genes (Basel). 2019. PMID: 31671674 Free PMC article. Review.
-
The MRE11 complex: A versatile toolkit for the repair of broken DNA.DNA Repair (Amst). 2020 Jul-Aug;91-92:102869. doi: 10.1016/j.dnarep.2020.102869. Epub 2020 May 15. DNA Repair (Amst). 2020. PMID: 32480356 Review.
Cited by
-
Characterization of the conserved regions of E1A protein from human adenovirus for reinforcement of cytotoxic T lymphocytes responses to the all genogroups causes ocular manifestation through an in silico approach.Iran J Microbiol. 2022 Oct;14(5):746-758. doi: 10.18502/ijm.v14i5.10971. Iran J Microbiol. 2022. PMID: 36531810 Free PMC article.
-
Post-Translational Modification of MRE11: Its Implication in DDR and Diseases.Genes (Basel). 2021 Jul 28;12(8):1158. doi: 10.3390/genes12081158. Genes (Basel). 2021. PMID: 34440334 Free PMC article. Review.
-
En Guard! The Interactions between Adenoviruses and the DNA Damage Response.Viruses. 2020 Sep 7;12(9):996. doi: 10.3390/v12090996. Viruses. 2020. PMID: 32906746 Free PMC article. Review.
References
-
- Ward JF, The yield of DNA double-strand breaks produced intracellularly by ionizing radiation: a review, Int. J. Radiat. Biol. 57 (1990) 1141–1150. - PubMed
-
- Stanulla M, Wang J, Chervinsky DS, Aplan PD, Topoisomerase II inhibitors induce DNA double-strand breaks at a specific site within the AML1 locus, Leukemia 11 (1997) 490–496. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
