

Comparing mutagenesis and simulations as tools for identifying functionally important sequence changes for protein thermal adaptation
- PMID: 30584112
- PMCID: PMC6329985
- DOI: 10.1073/pnas.1817455116
Comparing mutagenesis and simulations as tools for identifying functionally important sequence changes for protein thermal adaptation
Abstract
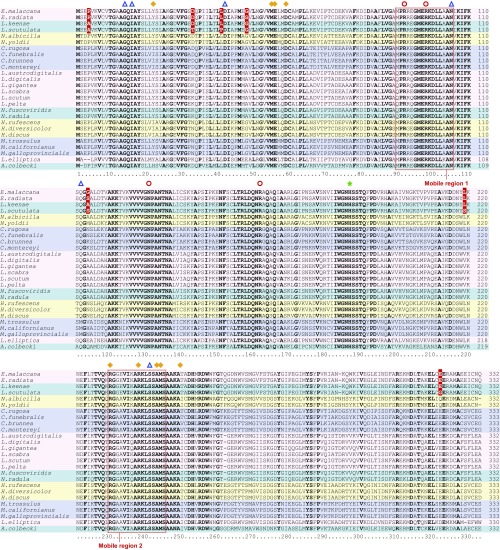
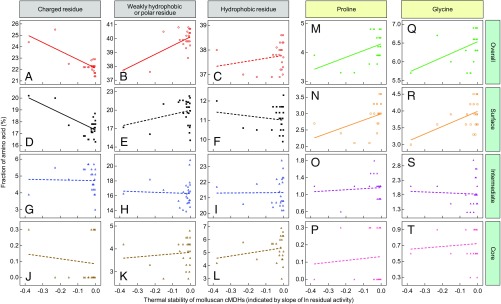
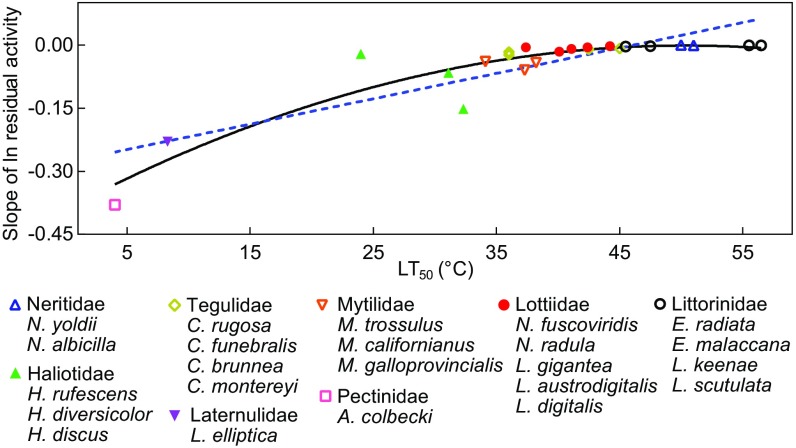
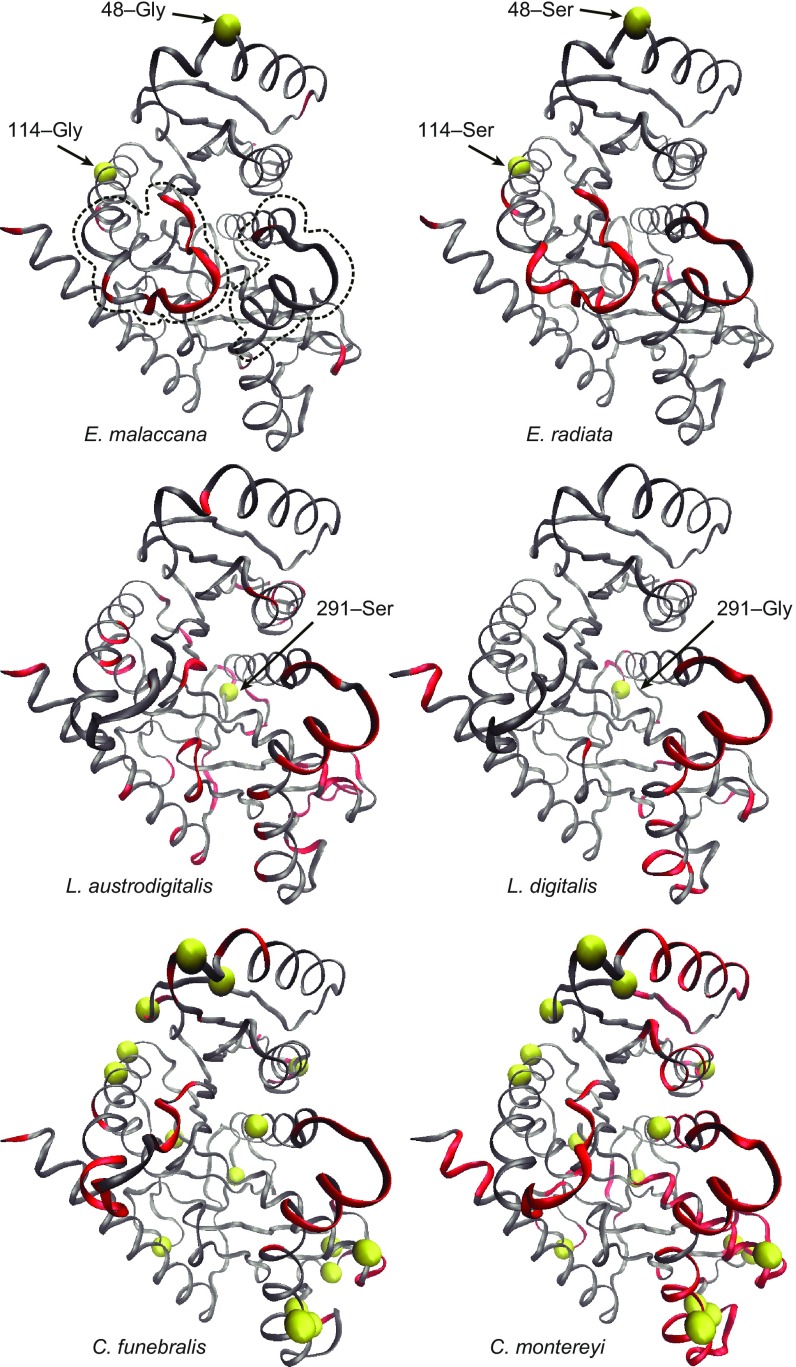
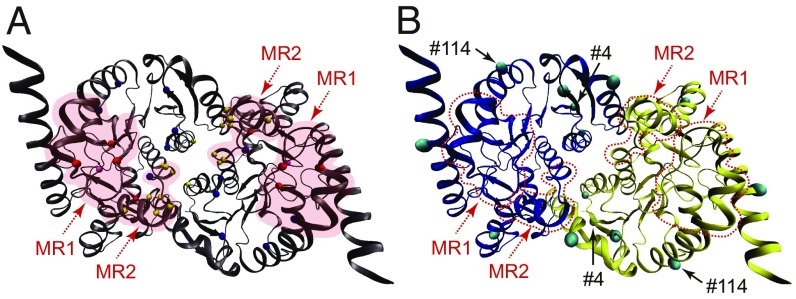
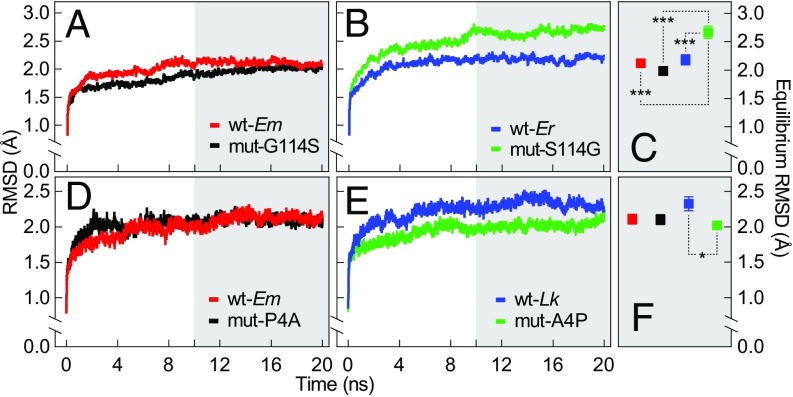
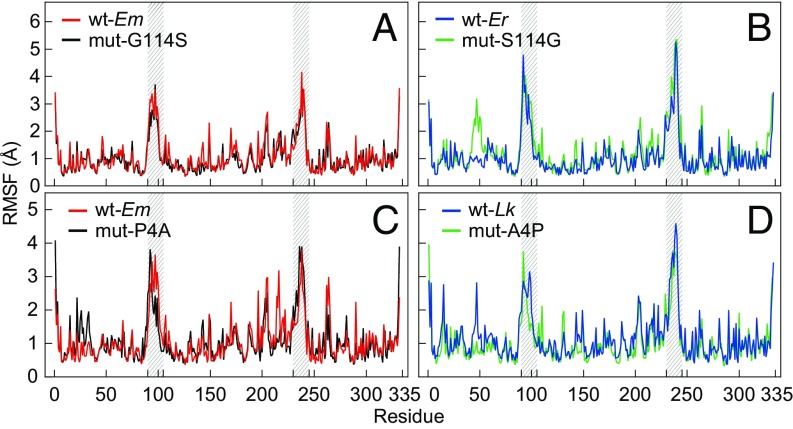
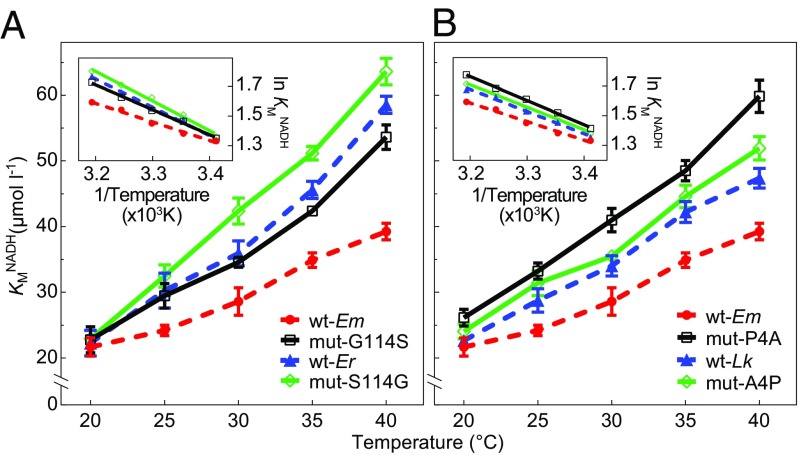
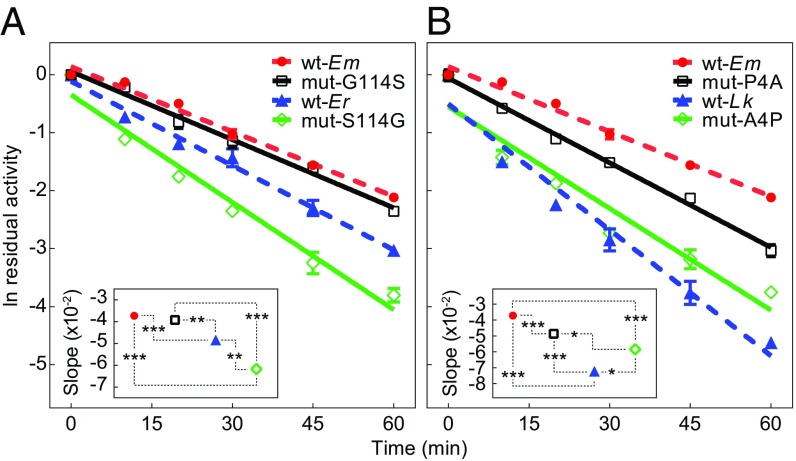
Comparative studies of orthologous proteins of species evolved at different temperatures have revealed consistent patterns of temperature-related variation in thermal stabilities of structure and function. However, the precise mechanisms by which interspecific variations in sequence foster these adaptive changes remain largely unknown. Here, we compare orthologs of cytosolic malate dehydrogenase (cMDH) from marine molluscs adapted to temperatures ranging from -1.9 °C (Antarctica) to ∼55 °C (South China coast) and show how amino acid usage in different regions of the enzyme (surface, intermediate depth, and protein core) varies with adaptation temperature. This eukaryotic enzyme follows some but not all of the rules established in comparisons of archaeal and bacterial proteins. To link the effects of specific amino acid substitutions with adaptive variations in enzyme thermal stability, we combined site-directed mutagenesis (SDM) and in vitro protein experimentation with in silico mutagenesis using molecular dynamics simulation (MDS) techniques. SDM and MDS methods generally but not invariably yielded common effects on protein stability. MDS analysis is shown to provide insights into how specific amino acid substitutions affect the conformational flexibilities of mobile regions (MRs) of the enzyme that are essential for binding and catalysis. Whereas these substitutions invariably lie outside of the MRs, they effectively transmit their flexibility-modulating effects to the MRs through linked interactions among surface residues. This discovery illustrates that regions of the protein surface lying outside of the site of catalysis can help establish an enzyme's thermal responses and foster evolutionary adaptation of function.
Keywords: adaptation; cytosolic malate dehydrogenase; evolution; molecular dynamics simulations; protein evolution.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Structural flexibility and protein adaptation to temperature: Molecular dynamics analysis of malate dehydrogenases of marine molluscs.Proc Natl Acad Sci U S A. 2018 Feb 6;115(6):1274-1279. doi: 10.1073/pnas.1718910115. Epub 2018 Jan 22. Proc Natl Acad Sci U S A. 2018. PMID: 29358381 Free PMC article.
-
Thermal Adaptation of Cytosolic Malate Dehydrogenase Revealed by Deep Learning and Coevolutionary Analysis.J Chem Theory Comput. 2025 Mar 25;21(6):3277-3287. doi: 10.1021/acs.jctc.4c01774. Epub 2025 Mar 13. J Chem Theory Comput. 2025. PMID: 40079215 Free PMC article.
-
Temperature sensitivities of cytosolic malate dehydrogenases from native and invasive species of marine mussels (genus Mytilus): sequence-function linkages and correlations with biogeographic distribution.J Exp Biol. 2006 Feb;209(Pt 4):656-67. doi: 10.1242/jeb.02036. J Exp Biol. 2006. PMID: 16449560
-
Adaptations of protein structure and function to temperature: there is more than one way to 'skin a cat'.J Exp Biol. 2015 Jun;218(Pt 12):1801-11. doi: 10.1242/jeb.114298. J Exp Biol. 2015. PMID: 26085658 Review.
-
Malate dehydrogenase: a model for structure, evolution, and catalysis.Protein Sci. 1994 Oct;3(10):1883-8. doi: 10.1002/pro.5560031027. Protein Sci. 1994. PMID: 7849603 Free PMC article. Review.
Cited by
-
Tissue-specific temperature dependence of RNA editing levels in zebrafish.BMC Biol. 2023 Nov 20;21(1):262. doi: 10.1186/s12915-023-01738-4. BMC Biol. 2023. PMID: 37981664 Free PMC article.
-
Acetylome Analysis Reveals Population Differentiation of the Pacific Oyster Crassostrea gigas in Response to Heat Stress.Mar Biotechnol (NY). 2020 Apr;22(2):233-245. doi: 10.1007/s10126-020-09947-6. Epub 2020 Jan 29. Mar Biotechnol (NY). 2020. PMID: 31997089
-
RNA editing generates mRNA isoforms with distinct stabilities that may expand the thermal tolerance of mRNA and proteins in Mytilus species.Zool Res. 2025 May 18;46(3):527-537. doi: 10.24272/j.issn.2095-8137.2024.383. Zool Res. 2025. PMID: 40259733 Free PMC article.
-
Bioinformatic analysis of subfamily-specific regions in 3D-structures of homologs to study functional diversity and conformational plasticity in protein superfamilies.Comput Struct Biotechnol J. 2021 Feb 23;19:1302-1311. doi: 10.1016/j.csbj.2021.02.005. eCollection 2021. Comput Struct Biotechnol J. 2021. PMID: 33738079 Free PMC article.
-
Principles of Cold Adaptation of Fish Lactate Dehydrogenases Revealed by Computer Simulations of the Catalytic Reaction.Mol Biol Evol. 2023 May 2;40(5):msad099. doi: 10.1093/molbev/msad099. Mol Biol Evol. 2023. PMID: 37116207 Free PMC article.
References
-
- Somero GN, Lockwood BL, Tomanek L. Biochemical Adaptation: Response to Environmental Challenges from Life’s Origins to the Anthropocene. Sinauer Associates; Sunderland, MA: 2017.
-
- Liao ML, et al. Heat-resistant cytosolic malate dehydrogenases (cMDHs) of thermophilic intertidal snails (genus Echinolittorina): Protein underpinnings of tolerance to body temperatures reaching 55°C. J Exp Biol. 2017;220:2066–2075. - PubMed
-
- Fields PA, Dong Y, Meng X, Somero GN. Adaptations of protein structure and function to temperature: There is more than one way to ‘skin a cat’. J Exp Biol. 2015;218:1801–1811. - PubMed
-
- Dalhus B, et al. Structural basis for thermophilic protein stability: Structures of thermophilic and mesophilic malate dehydrogenases. J Mol Biol. 2002;318:707–721. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
