

High affinity binding of H3K14ac through collaboration of bromodomains 2, 4 and 5 is critical for the molecular and tumor suppressor functions of PBRM1
- PMID: 30585695
- PMCID: PMC6441893
- DOI: 10.1002/1878-0261.12434
High affinity binding of H3K14ac through collaboration of bromodomains 2, 4 and 5 is critical for the molecular and tumor suppressor functions of PBRM1
Abstract
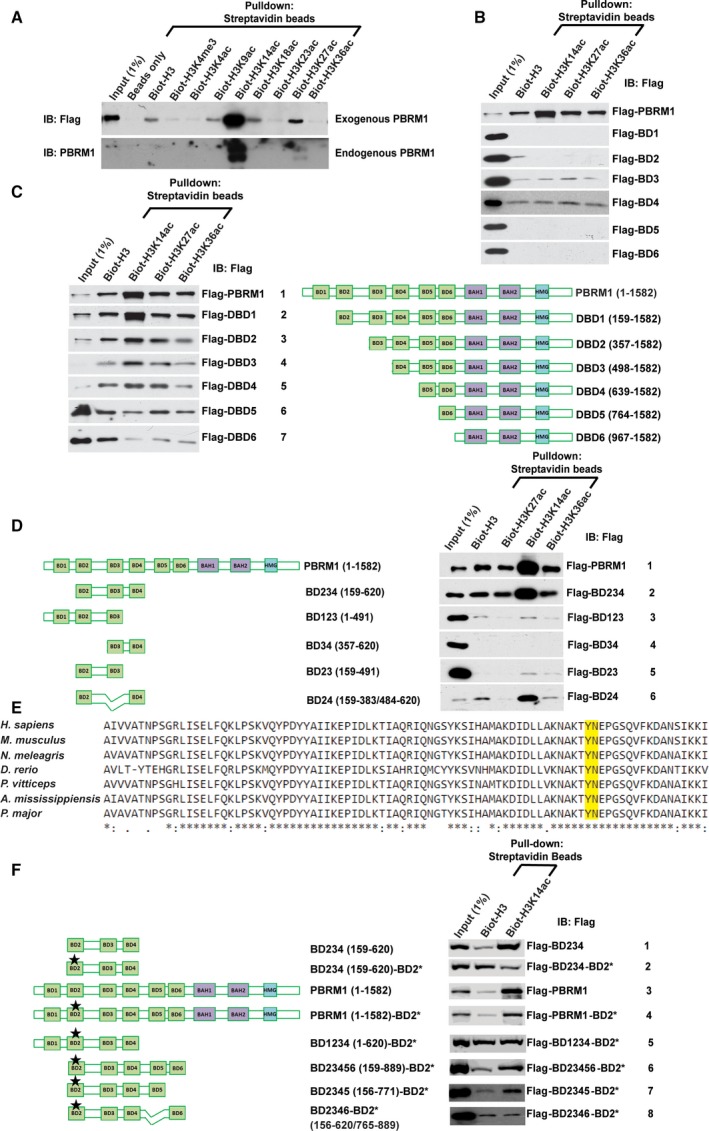
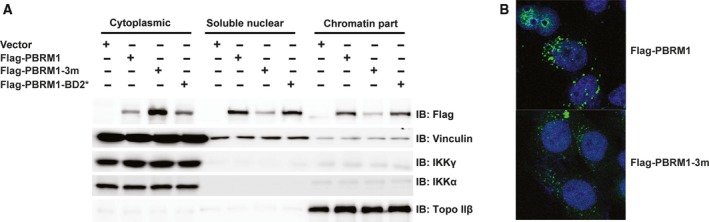
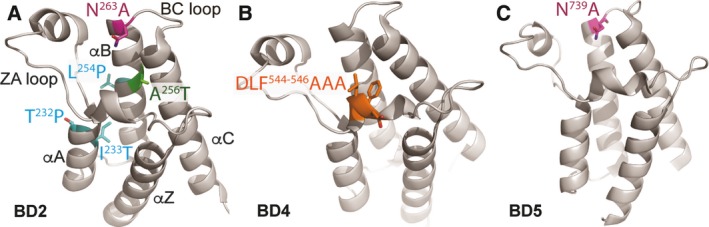
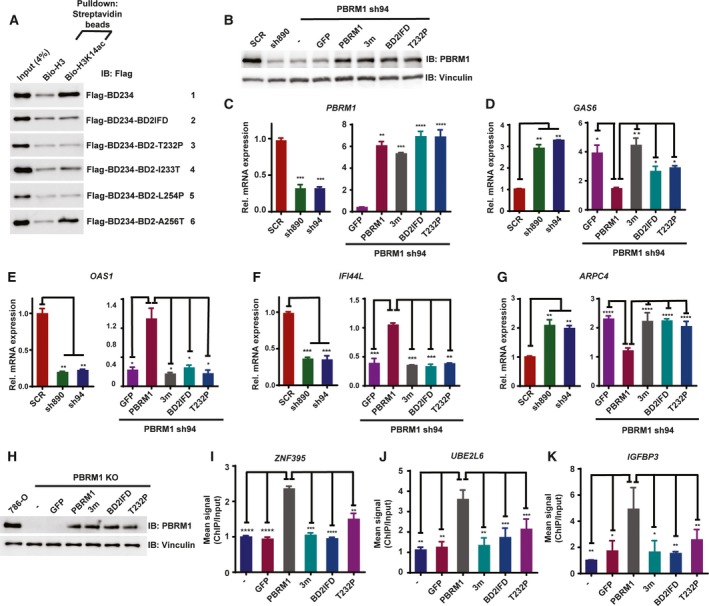
Polybromo-1 (PBRM1) is an important tumor suppressor in kidney cancer. It contains six tandem bromodomains (BDs), which are specialized structures that recognize acetyl-lysine residues. While BD2 has been found to bind acetylated histone H3 lysine 14 (H3K14ac), it is not known whether other BDs collaborate with BD2 to generate strong binding to H3K14ac, and the importance of H3K14ac recognition for the molecular and tumor suppressor function of PBRM1 is also unknown. We discovered that full-length PBRM1, but not its individual BDs, strongly binds H3K14ac. BDs 2, 4, and 5 were found to collaborate to facilitate strong binding to H3K14ac. Quantitative measurement of the interactions between purified BD proteins and H3K14ac or nonacetylated peptides confirmed the tight and specific association of the former. Interestingly, while the structural integrity of BD4 was found to be required for H3K14ac recognition, the conserved acetyl-lysine binding site of BD4 was not. Furthermore, simultaneous point mutations in BDs 2, 4, and 5 prevented recognition of H3K14ac, altered promoter binding and gene expression, and caused PBRM1 to relocalize to the cytoplasm. In contrast, tumor-derived point mutations in BD2 alone lowered PBRM1's affinity to H3K14ac and also disrupted promoter binding and gene expression without altering cellular localization. Finally, overexpression of PBRM1 variants containing point mutations in BDs 2, 4, and 5 or BD2 alone failed to suppress tumor growth in a xenograft model. Taken together, our study demonstrates that BDs 2, 4, and 5 of PBRM1 collaborate to generate high affinity to H3K14ac and tether PBRM1 to chromatin. Mutations in BD2 alone weaken these interactions, and this is sufficient to abolish its molecular and tumor suppressor functions.
Keywords: H3K14ac; PBRM1; bromodomain; kidney cancer; synergy.
© 2018 The Authors. Published by FEBS Press and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interests.
Figures



References
-
- Benusiglio PR, Couve S, Gilbert‐Dussardier B, Deveaux S, Le Jeune H, Da Costa M, Fromont G, Memeteau F, Yacoub M, Coupier I et al (2015) A germline mutation in PBRM1 predisposes to renal cell carcinoma. J Med Genet 52, 426–430. - PubMed
-
- Chandrasekaran R and Thompson M (2007) Polybromo‐1‐bromodomains bind histone H3 at specific acetyl‐lysine positions. Biochem Biophys Res Commun 355, 661–666. - PubMed
-
- Choueiri TK and Motzer RJ (2017) Systemic Therapy for Metastatic Renal‐Cell Carcinoma. N Engl J Med 376, 354–366. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
