

Ate1-mediated posttranslational arginylation affects substrate adhesion and cell migration in Dictyostelium discoideum
- PMID: 30586322
- PMCID: PMC6594445
- DOI: 10.1091/mbc.E18-02-0132
Ate1-mediated posttranslational arginylation affects substrate adhesion and cell migration in Dictyostelium discoideum
Abstract
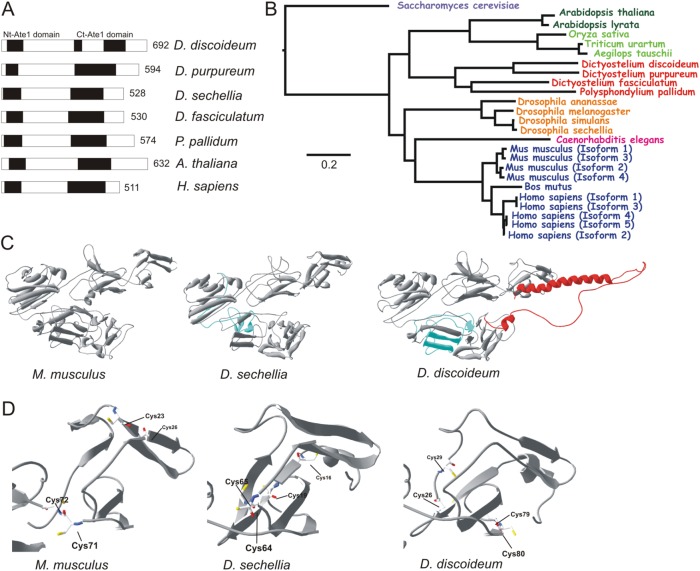
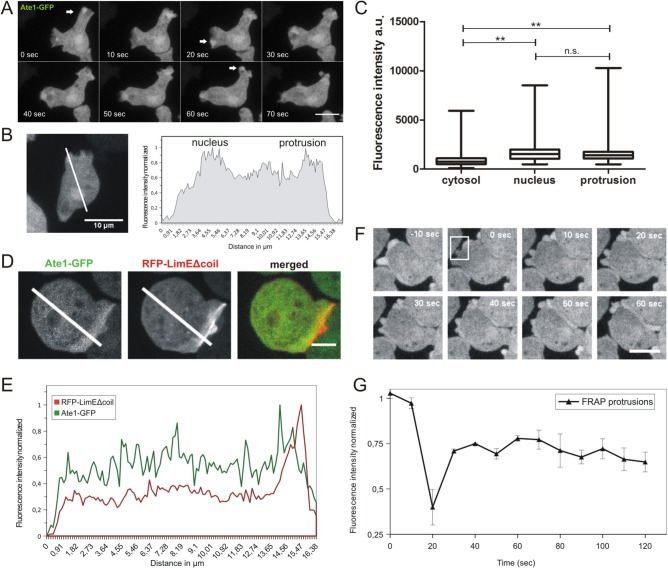
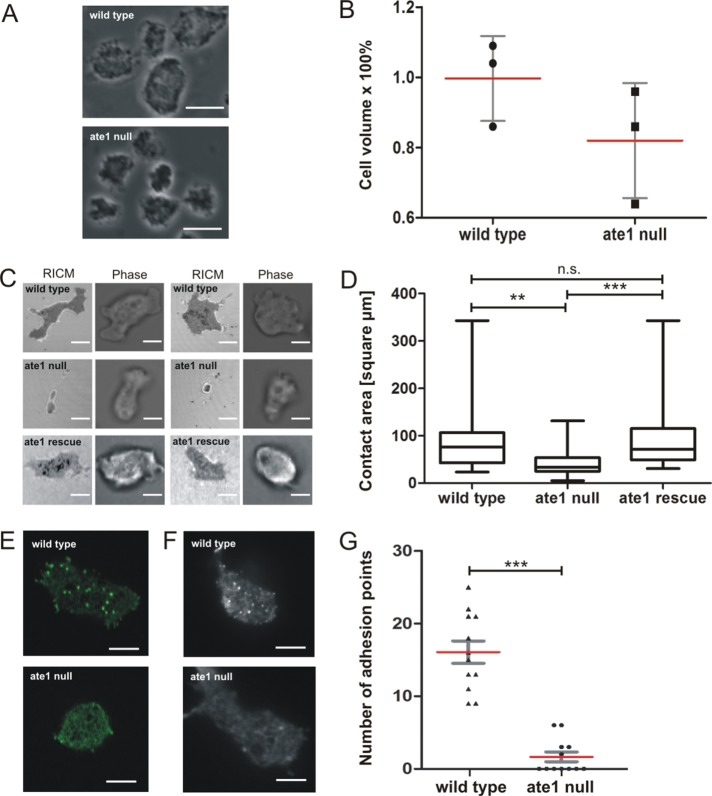
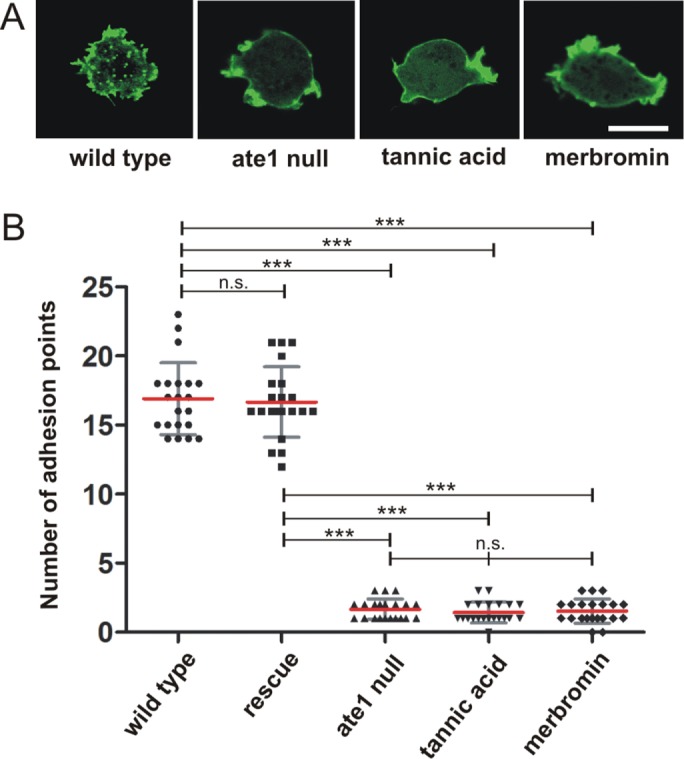
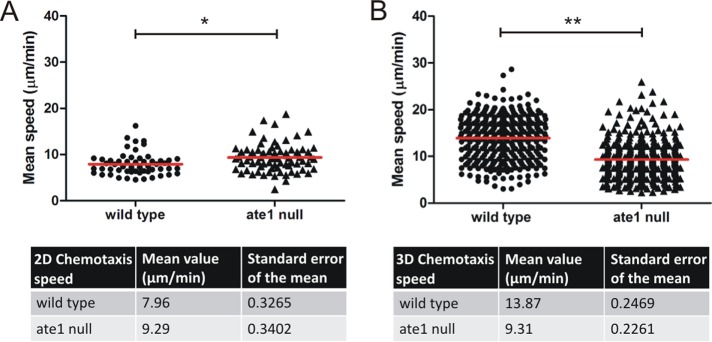
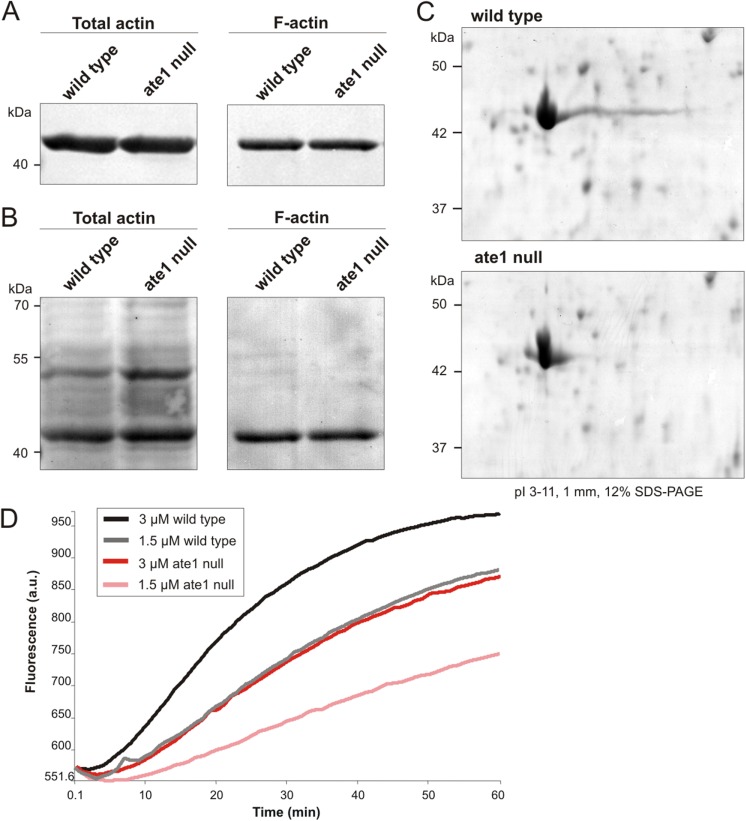
The highly conserved enzyme arginyl-tRNA-protein transferase (Ate1) mediates arginylation, a posttranslational modification that is only incompletely understood at its molecular level. To investigate whether arginylation affects actin-dependent processes in a simple model organism, Dictyostelium discoideum, we knocked out the gene encoding Ate1 and characterized the phenotype of ate1-null cells. Visualization of actin cytoskeleton dynamics by live-cell microscopy indicated significant changes in comparison to wild-type cells. Ate1-null cells were almost completely lacking focal actin adhesion sites at the substrate-attached surface and were only weakly adhesive. In two-dimensional chemotaxis assays toward folate or cAMP, the motility of ate1-null cells was increased. However, in three-dimensional chemotaxis involving more confined conditions, the motility of ate1-null cells was significantly reduced. Live-cell imaging showed that GFP-tagged Ate1 rapidly relocates to sites of newly formed actin-rich protrusions. By mass spectrometric analysis, we identified four arginylation sites in the most abundant actin isoform of Dictyostelium, in addition to arginylation sites in other actin isoforms and several actin-binding proteins. In vitro polymerization assays with actin purified from ate1-null cells revealed a diminished polymerization capacity in comparison to wild-type actin. Our data indicate that arginylation plays a crucial role in the regulation of cytoskeletal activities.
Figures

References
-
- Arnold K, Bordoli L, Kopp J, Schwede T. (2006). The SWISS-MODEL workspace: a web-based environment for protein structure homology modelling. Bioinformatics , 195–201. - PubMed
-
- Axelrod D. (2003). Total internal reflection fluorescence microscopy in cell biology. Methods Enzymol , 1–33. - PubMed
-
- Balzi E, Choder M, Chen WN, Varshavsky A, Goffeau A. (1990). Cloning and functional analysis of the arginyl-tRNA-protein transferase gene ATE1 of Saccharomyces cerevisiae. J Biol Chem , 7464–7471. - PubMed
-
- Bohley P, Kopitz J, Adam G, Rist B, von Appen F, Urban S. (1991). Post-translational arginylation and intracellular proteolysis. Biomed Biochim Acta , 343–346. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
