

LMO7 Is a Negative Feedback Regulator of Transforming Growth Factor β Signaling and Fibrosis
- PMID: 30586711
- PMCID: PMC6371979
- DOI: 10.1161/CIRCULATIONAHA.118.034615
LMO7 Is a Negative Feedback Regulator of Transforming Growth Factor β Signaling and Fibrosis
Abstract
Background: Vascular smooth muscle cells (SMCs) synthesize extracellular matrix (ECM) that contributes to tissue remodeling after revascularization interventions. The cytokine transforming growth factor β (TGF-β) is induced on tissue injury and regulates tissue remodeling and wound healing, but dysregulated signaling results in excess ECM deposition and fibrosis. The LIM (Lin11, Isl-1 & Mec-3) domain protein LIM domain only 7 (LMO7) is a TGF-β1 target gene in hepatoma cells, but its role in vascular physiology and fibrosis is unknown.
Methods: We use carotid ligation and femoral artery denudation models in mice with global or inducible smooth muscle-specific deletion of LMO7, and knockout, knockdown, overexpression, and mutagenesis approaches in mouse and human SMC, and human arteriovenous fistula and cardiac allograft vasculopathy samples to assess the role of LMO7 in neointima and fibrosis.
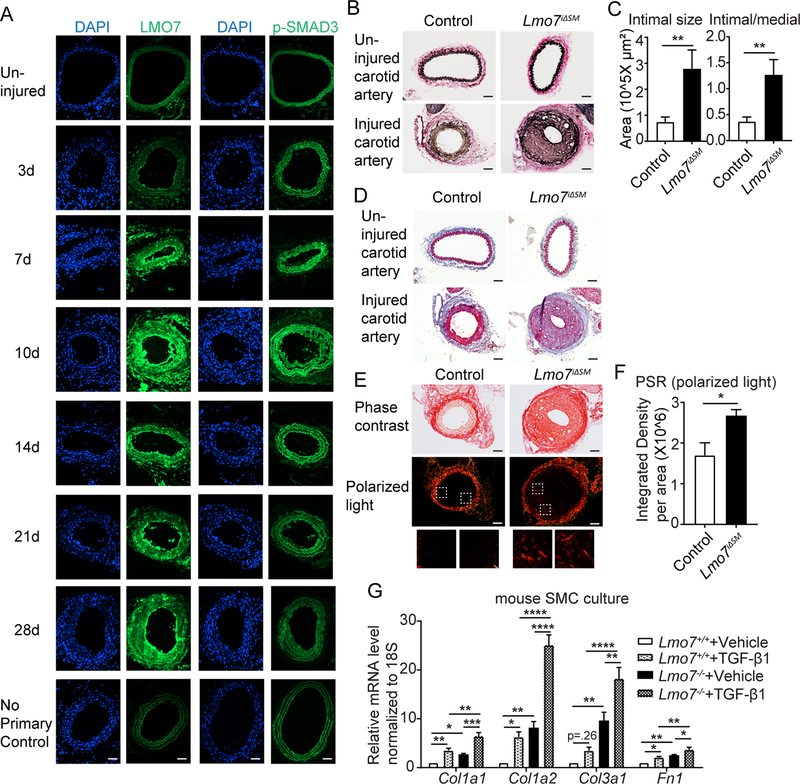
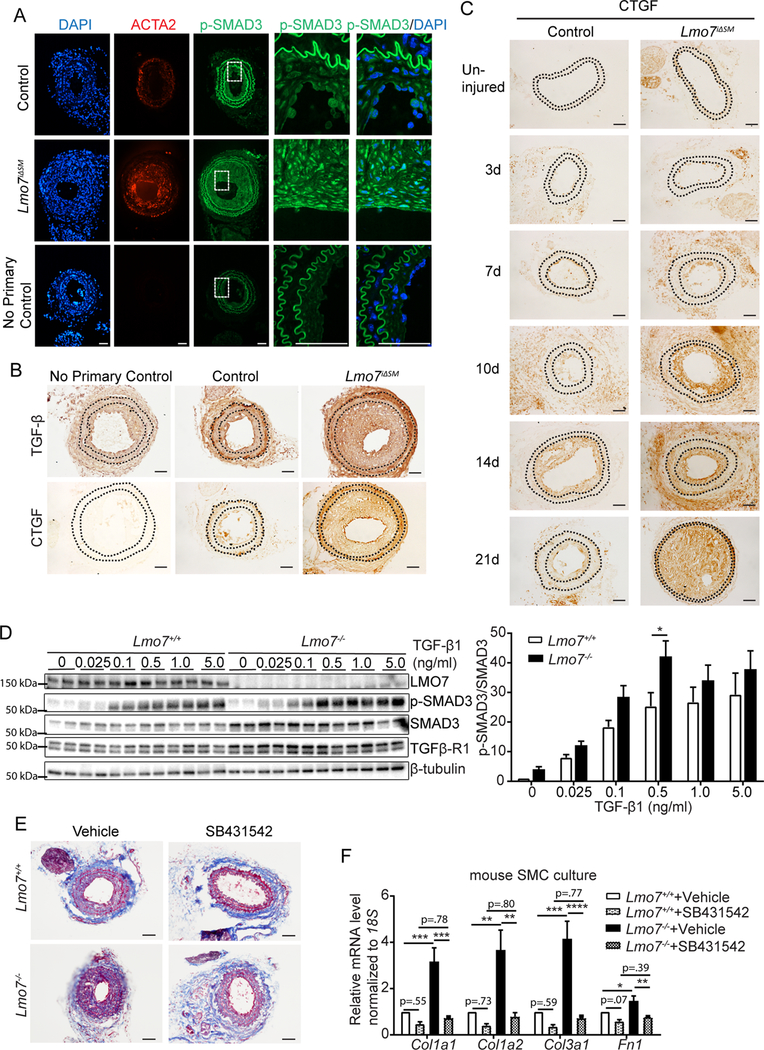
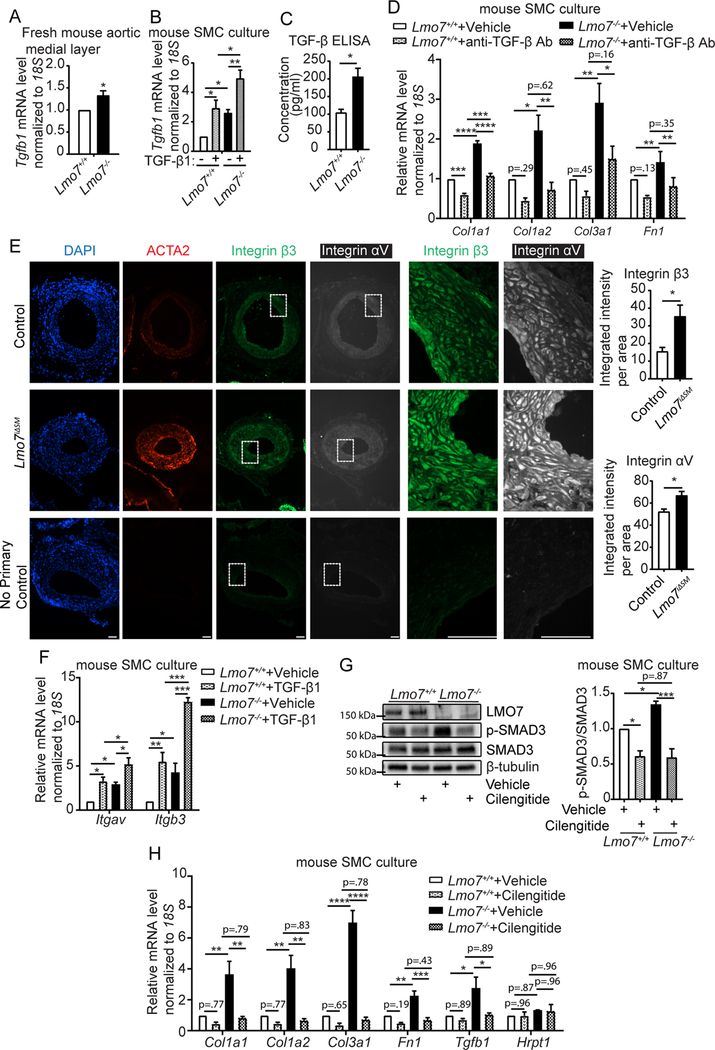
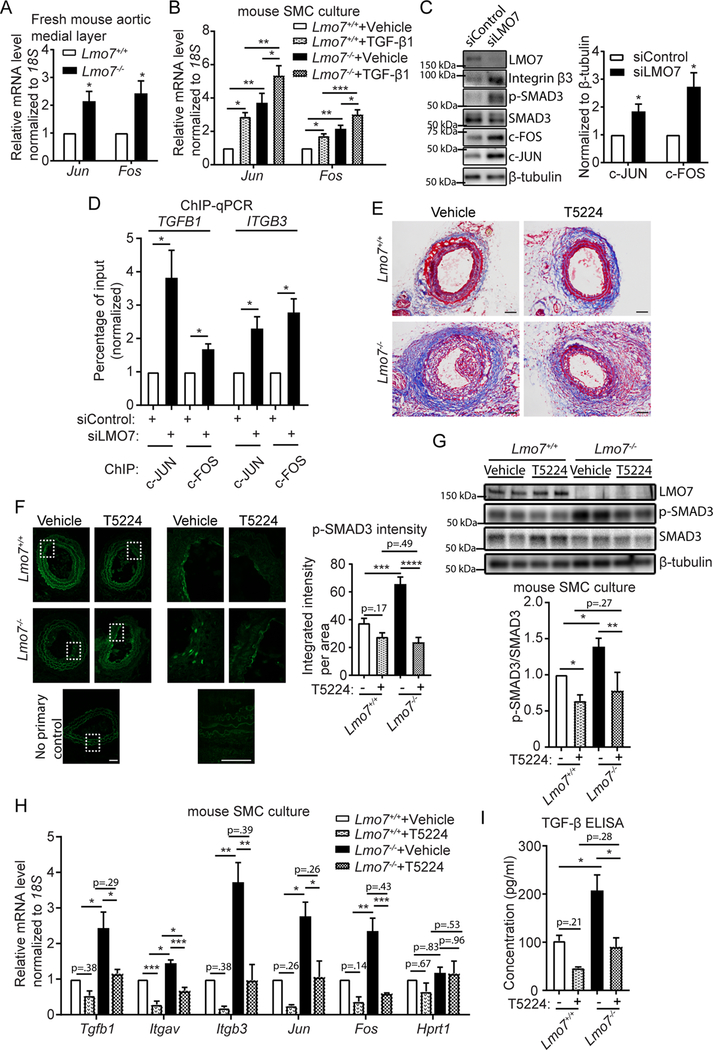
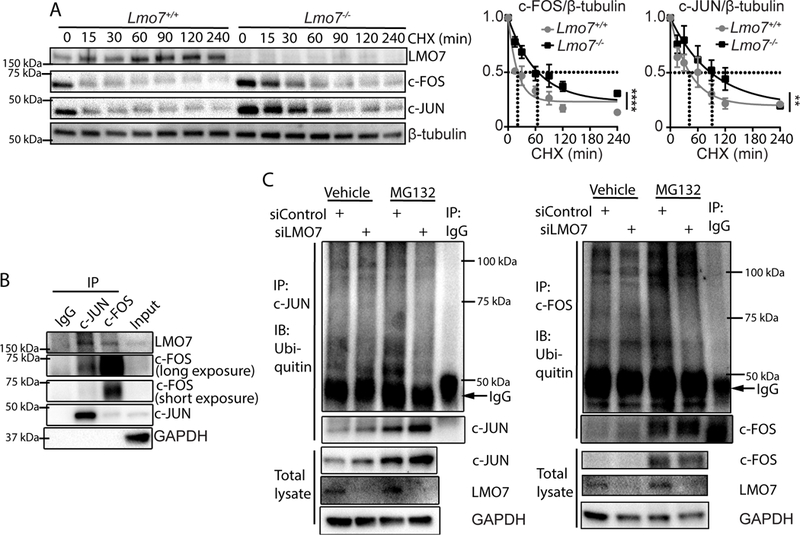
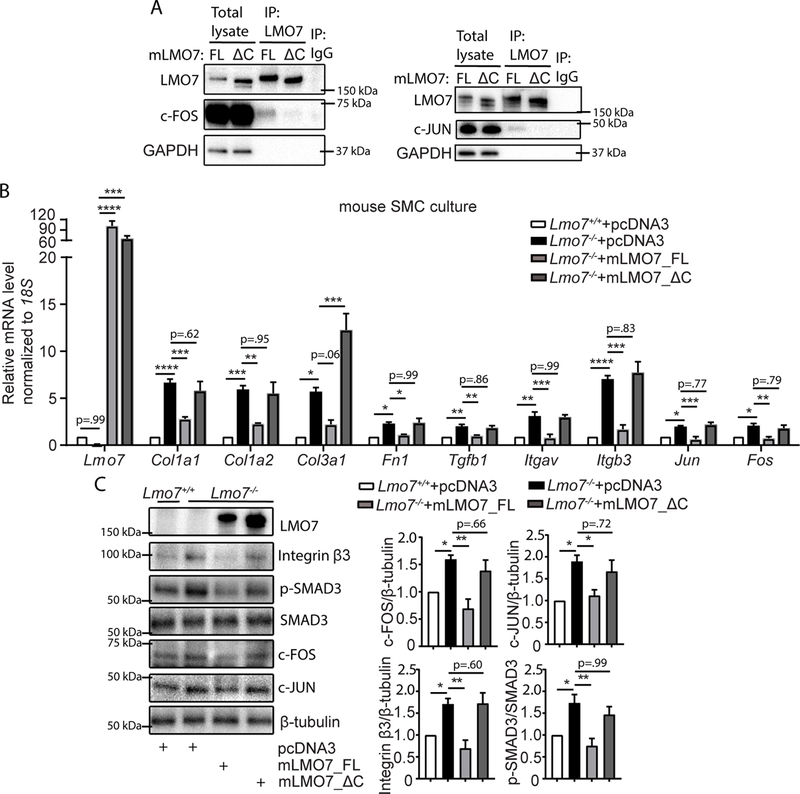
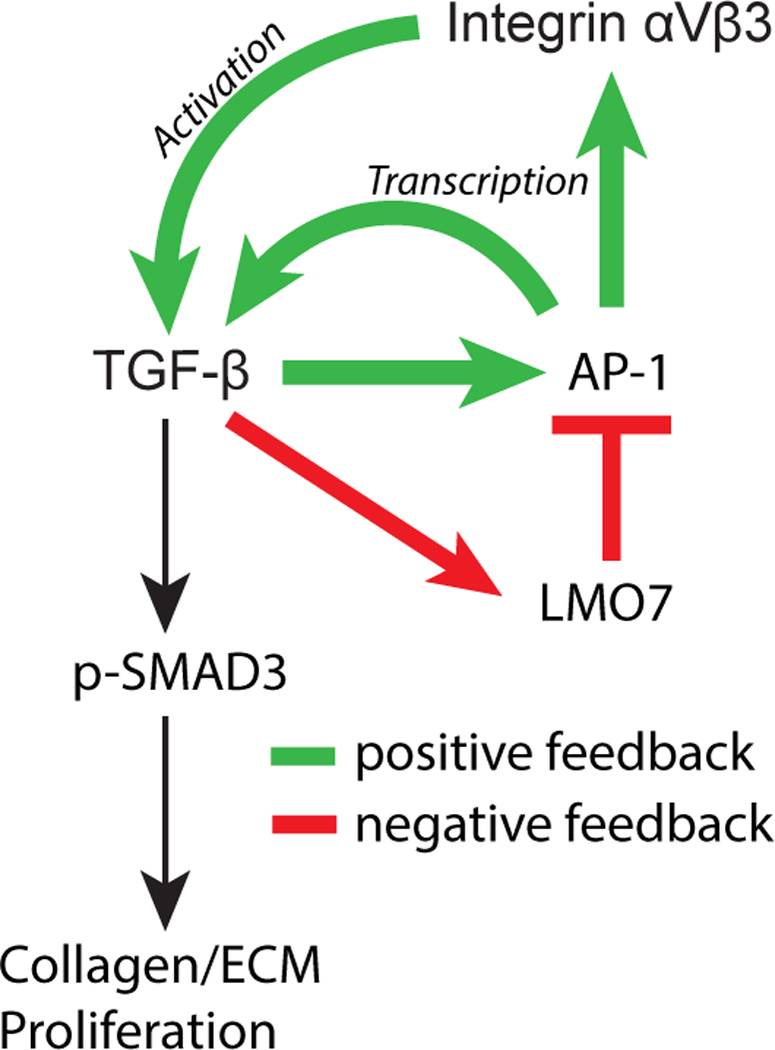
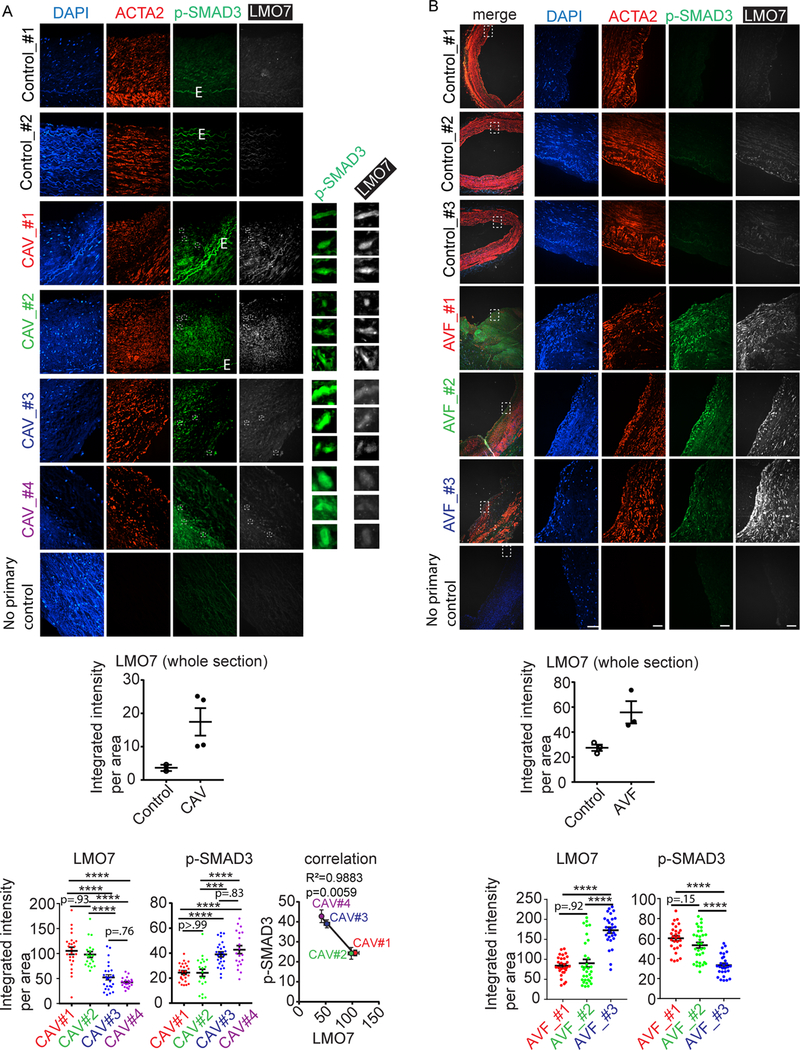
Results: We demonstrate that LMO7 is induced postinjury and by TGF-β in SMC in vitro. Global or SMC-specific LMO7 deletion enhanced neointimal formation, TGF-β signaling, ECM deposition, and proliferation in vascular injury models. LMO7 loss of function in human and mouse SMC enhanced ECM protein expression at baseline and after TGF-β treatment. TGF-β neutralization or receptor antagonism prevented the exacerbated neointimal formation and ECM synthesis conferred by loss of LMO7. Notably, loss of LMO7 coordinately amplified TGF-β signaling by inducing expression of Tgfb1 mRNA, TGF-β protein, αv and β3 integrins that promote activation of latent TGF-β, and downstream effectors SMAD3 phosphorylation and connective tissue growth factor. Mechanistically, the LMO7 LIM domain interacts with activator protein 1 transcription factor subunits c-FOS and c-JUN and promotes their ubiquitination and degradation, disrupting activator protein 1-dependent TGF-β autoinduction. Importantly, preliminary studies suggest that LMO7 is upregulated in human intimal hyperplastic arteriovenous fistula and cardiac allograft vasculopathy samples, and inversely correlates with SMAD3 phosphorylation in cardiac allograft vasculopathy.
Conclusions: LMO7 is induced by TGF-β and serves to limit vascular fibrotic responses through negative feedback regulation of the TGF-β pathway. This mechanism has important implications for intimal hyperplasia, wound healing, and fibrotic diseases.
Keywords: extracellular matrix; fibrosis; smooth muscle; transcription factors; transforming growth factor.
Figures
References
-
- Dangas GD, Claessen BE, Caixeta A, Sanidas EA, Mintz GS and Mehran R. In-stent restenosis in the drug-eluting stent era. J Am Coll Cardiol 2010;56:1897–1907. - PubMed
-
- Owens GK, Kumar MS and Wamhoff BR. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol Rev 2004;84:767–801. - PubMed
-
- Fattori R and Piva T. Drug-eluting stents in vascular intervention. Lancet 2003;361:247–249. - PubMed
-
- Leask A and Abraham DJ. TGF-beta signaling and the fibrotic response. FASEB J 2004;18:816–827. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL115247/HL/NHLBI NIH HHS/United States
- R01 HL118430/HL/NHLBI NIH HHS/United States
- R01 HL126933/HL/NHLBI NIH HHS/United States
- U54 HL117798/HL/NHLBI NIH HHS/United States
- R01 HL074190/HL/NHLBI NIH HHS/United States
- R01 HL122815/HL/NHLBI NIH HHS/United States
- R01 HL125850/HL/NHLBI NIH HHS/United States
- R01 HL153599/HL/NHLBI NIH HHS/United States
- R01 HL146101/HL/NHLBI NIH HHS/United States
- R01 HL142090/HL/NHLBI NIH HHS/United States
- R01 HL109233/HL/NHLBI NIH HHS/United States
- T32 GM007324/GM/NIGMS NIH HHS/United States
- R01 HL128406/HL/NHLBI NIH HHS/United States
- R56 HL117064/HL/NHLBI NIH HHS/United States
- P01 DK057751/DK/NIDDK NIH HHS/United States
- R01 HL091013/HL/NHLBI NIH HHS/United States
- R01 HL119529/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
