

Y-chromosomes can constrain adaptive evolution via epistatic interactions with other chromosomes
- PMID: 30587105
- PMCID: PMC6307184
- DOI: 10.1186/s12862-018-1327-6
Y-chromosomes can constrain adaptive evolution via epistatic interactions with other chromosomes
Abstract
Background: Variation in the non-coding regions of Y-chromosomes have been shown to influence gene regulation throughout the genome in some systems; a phenomenon termed Y-linked regulatory variation (YRV). This type of sex-specific genetic variance could have important implications for the evolution of male and female traits. If YRV contributes to the additive genetic variation of an autosomally coded trait shared between the sexes (e.g. body size), then selection could facilitate sexually dimorphic evolution via the Y-chromosome. In contrast, if YRV is entirely non-additive (i.e. interacts epistatically with other chromosomes), then Y-chromosomes could constrain trait evolution in both sexes whenever they are selected for the same trait value. The ability for this phenomenon to influence such fundamental evolutionary dynamics remains unexplored.
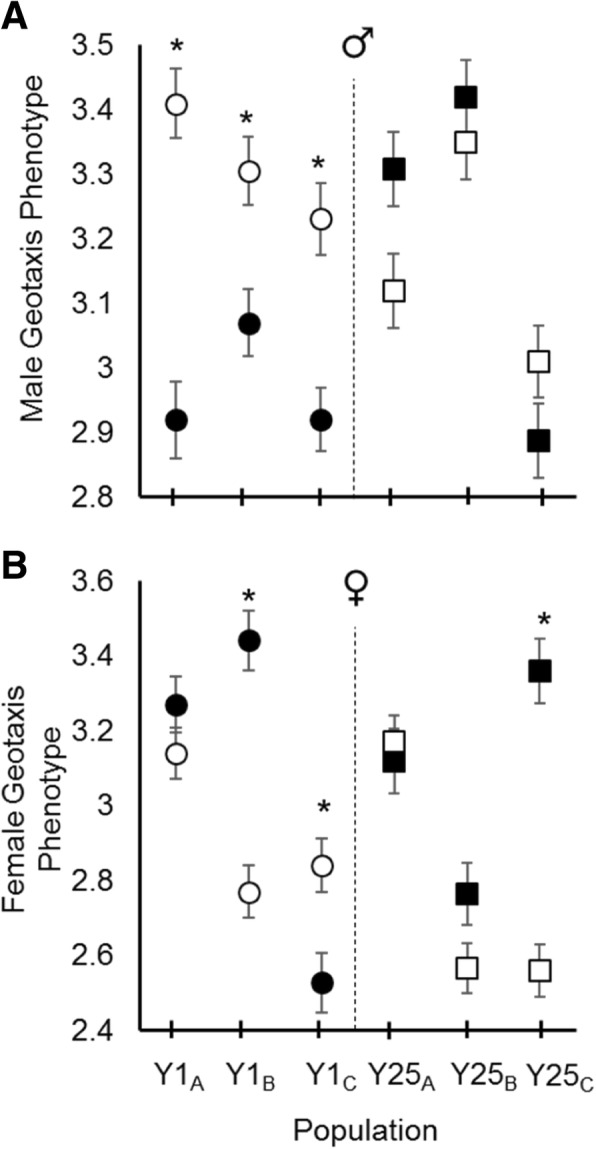
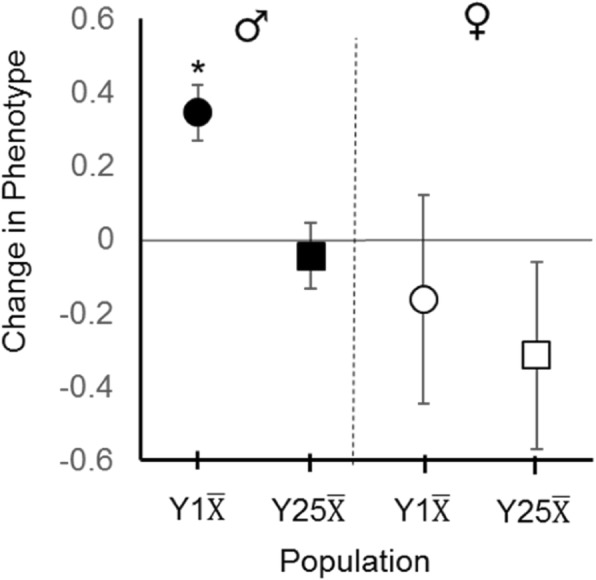
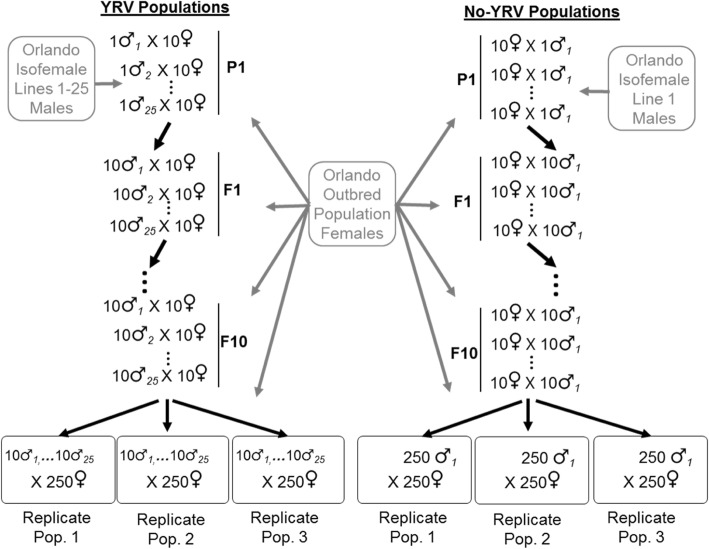
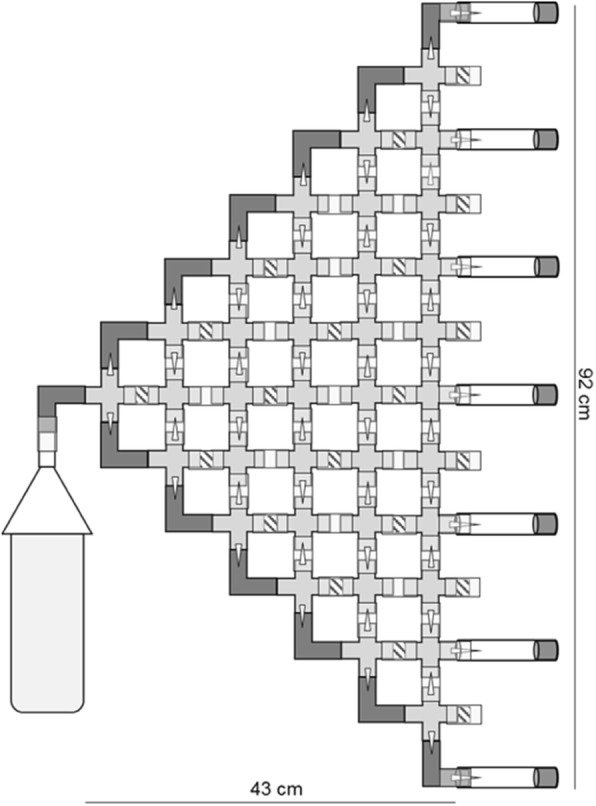
Results: Here we address the evolutionary contribution of Y-linked variance by selecting for improved male geotaxis in populations possessing multiple Y-chromosomes (i.e. possessed Y-linked additive and/or epistatic variation) or a single Y-chromosome variant (i.e. possessed no Y-linked variation). We found that males from populations possessing Y-linked variation did not significantly respond to selection; however, males from populations with no Y-linked variation did respond. These patterns suggest the presence of a large quantity of Y-linked epistatic variance in the multi-Y population that dramatically slowed its response.
Conclusions: Our results imply that YRV is unlikely to facilitate the evolution of sexually dimorphic traits (at least for the trait examined here), but can interfere with the rate of trait evolution in both males and females. This result could have real biological implications as it suggests that YRV can affect how quickly a population responds to new selective pressures (e.g. invasive species, novel pathogens, or climate change). Considering that YRV influences hundreds of genes and is likely typical of other independently-evolved hemizygous chromosomes, YRV-like phenomena may represent common and significant costs to hemizygous sex determination.
Keywords: Drosophila melanogaster; Epistasis; Evolutionary constraint; Selection; Y-linked regulatory variation.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
Similar articles
-
A test for Y-linked additive and epistatic effects on surviving bacterial infections in Drosophila melanogaster.J Evol Biol. 2017 Jul;30(7):1400-1408. doi: 10.1111/jeb.13118. Epub 2017 Jun 14. J Evol Biol. 2017. PMID: 28510293
-
Gene-poor Y-chromosomes substantially impact male trait heritabilities and may help shape sexually dimorphic evolution.Heredity (Edinb). 2023 Apr;130(4):236-241. doi: 10.1038/s41437-023-00596-8. Epub 2023 Feb 10. Heredity (Edinb). 2023. PMID: 36759734 Free PMC article.
-
Y-linked variation for autosomal immune gene regulation has the potential to shape sexually dimorphic immunity.Proc Biol Sci. 2015 Dec 7;282(1820):20151301. doi: 10.1098/rspb.2015.1301. Proc Biol Sci. 2015. PMID: 26631557 Free PMC article.
-
Epistasis for quantitative traits in Drosophila.Methods Mol Biol. 2015;1253:47-70. doi: 10.1007/978-1-4939-2155-3_4. Methods Mol Biol. 2015. PMID: 25403527 Review.
-
The evolution of hybrid infertility: perpetual coevolution between gender-specific and sexually antagonistic genes.Genetica. 2002 Nov;116(2-3):179-88. Genetica. 2002. PMID: 12555776 Review.
Cited by
-
State of the art on the physical mapping of the Y-chromosome in the Bovidae and comparison with other species - A review.Anim Biosci. 2022 Sep;35(9):1289-1302. doi: 10.5713/ab.21.0480. Epub 2022 Mar 2. Anim Biosci. 2022. PMID: 35240029 Free PMC article.
-
Y chromosome-linked variation affects locomotor activity in male Drosophila melanogaster and is robust to differences in thermal environment.Heredity (Edinb). 2023 May;130(5):312-319. doi: 10.1038/s41437-023-00604-x. Epub 2023 Mar 13. Heredity (Edinb). 2023. PMID: 36914794 Free PMC article.
-
Who's your daddy? Behavioral and epigenetic consequences of paternal drug exposure.Int J Dev Neurosci. 2019 Nov;78:109-121. doi: 10.1016/j.ijdevneu.2019.07.002. Epub 2019 Jul 10. Int J Dev Neurosci. 2019. PMID: 31301337 Free PMC article. Review.
References
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
