

Genetic Analysis of the Organization, Development, and Plasticity of Corneal Innervation in Mice
- PMID: 30587537
- PMCID: PMC6381234
- DOI: 10.1523/JNEUROSCI.1401-18.2018
Genetic Analysis of the Organization, Development, and Plasticity of Corneal Innervation in Mice
Abstract
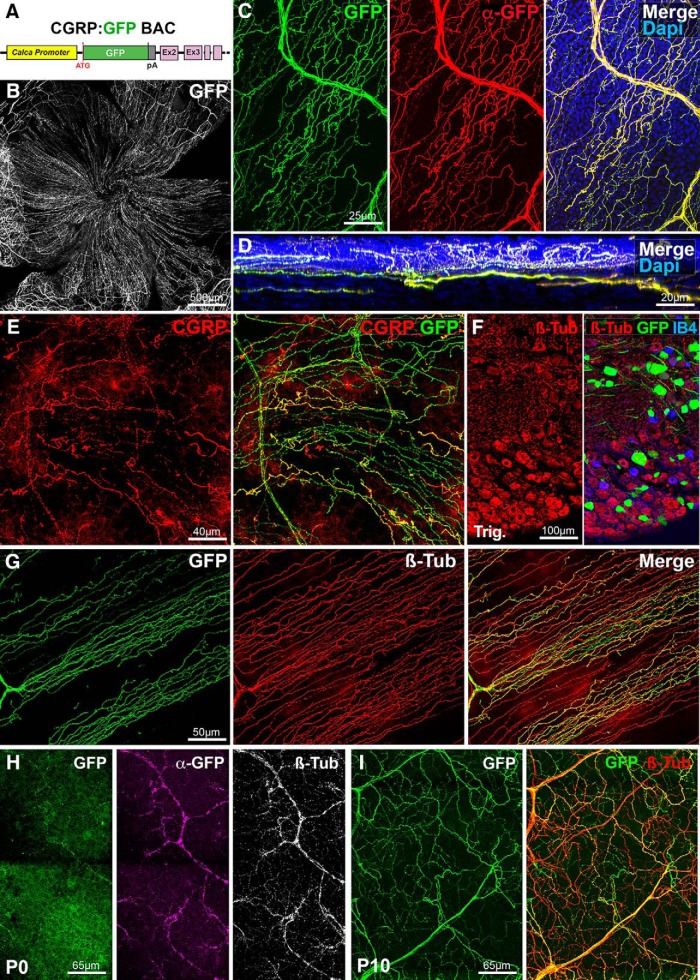
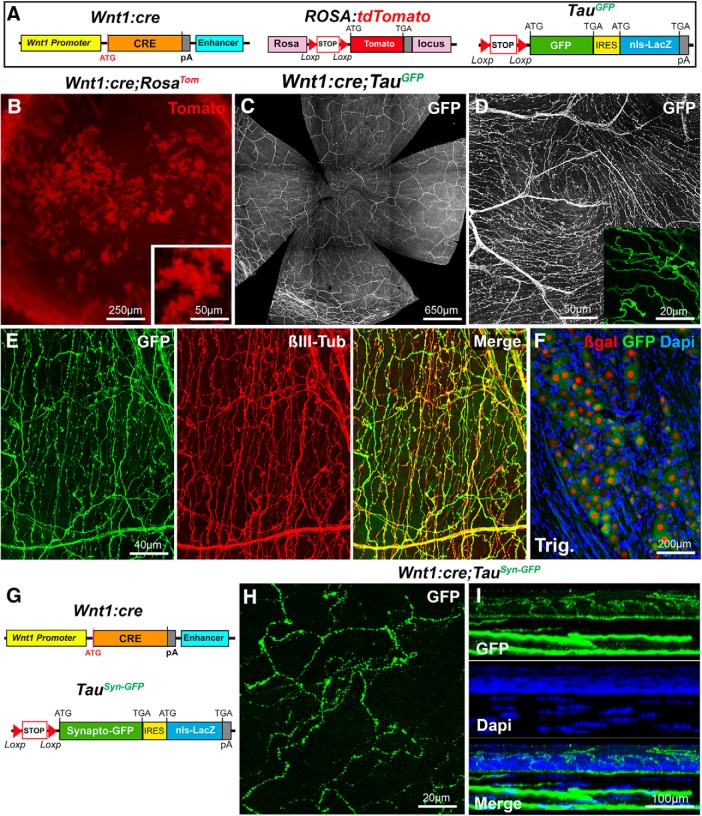
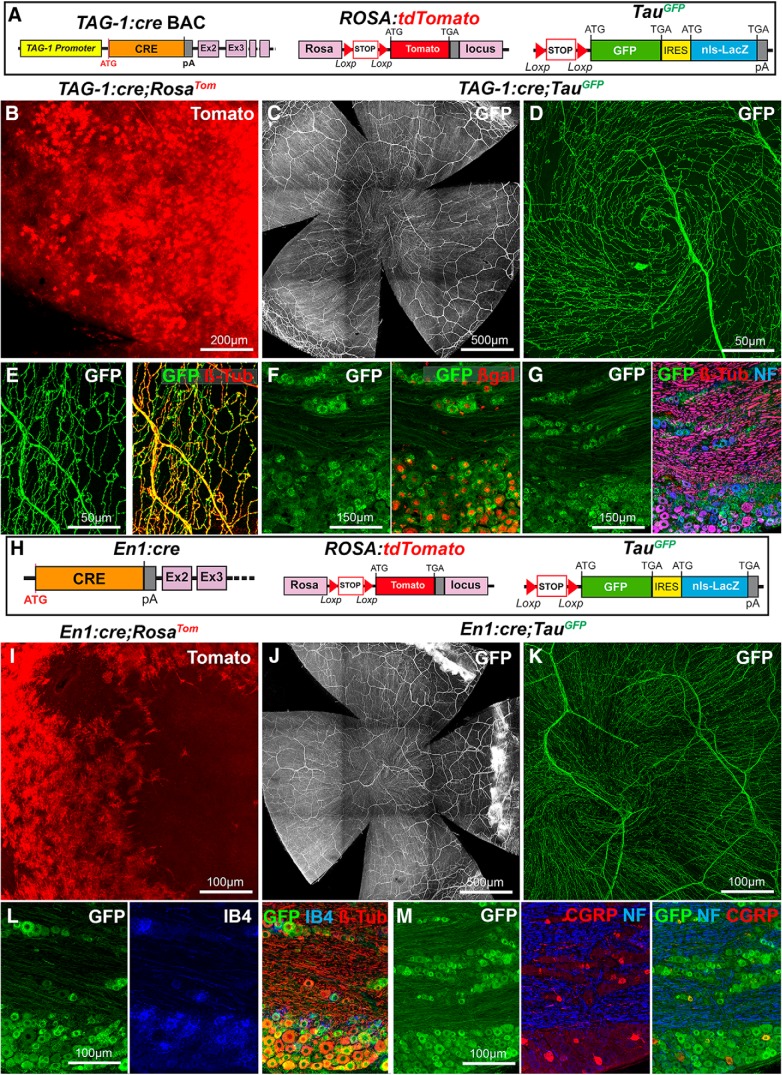
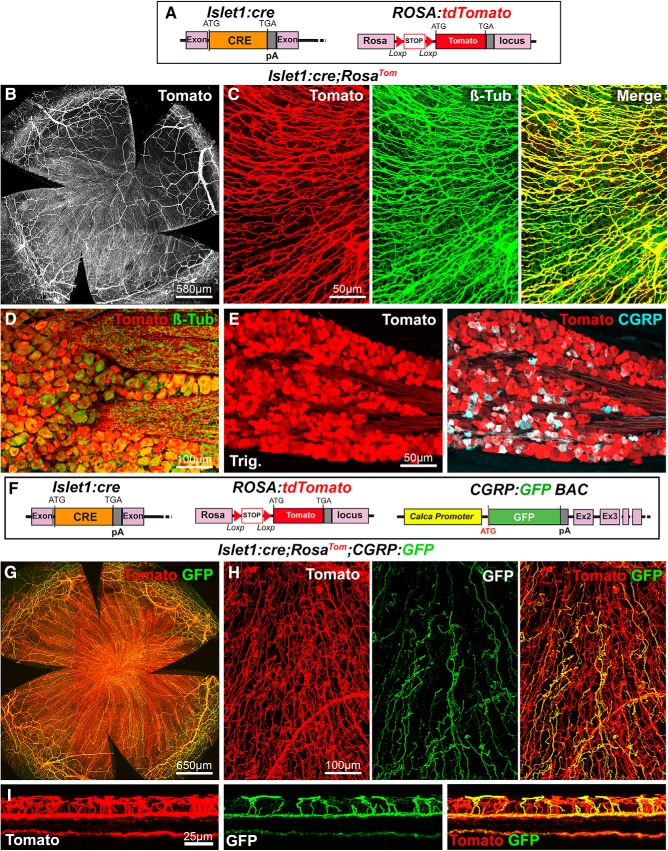
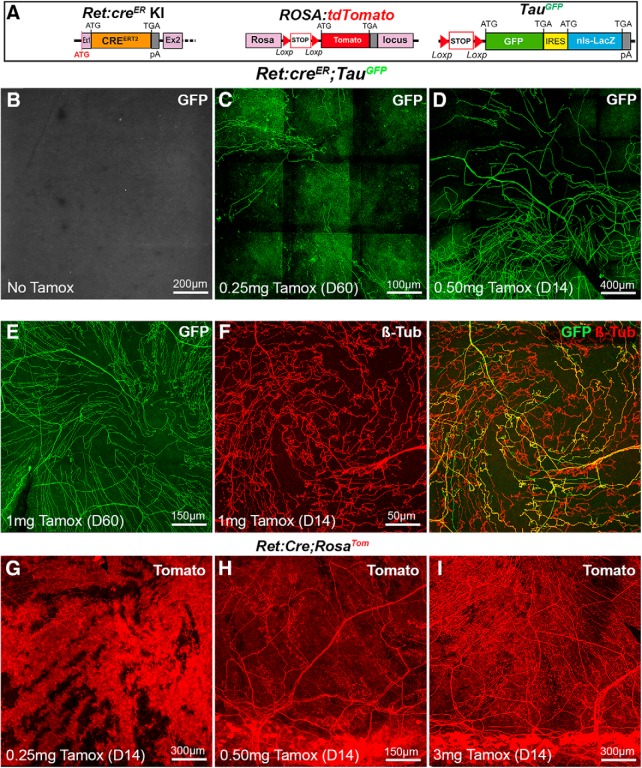
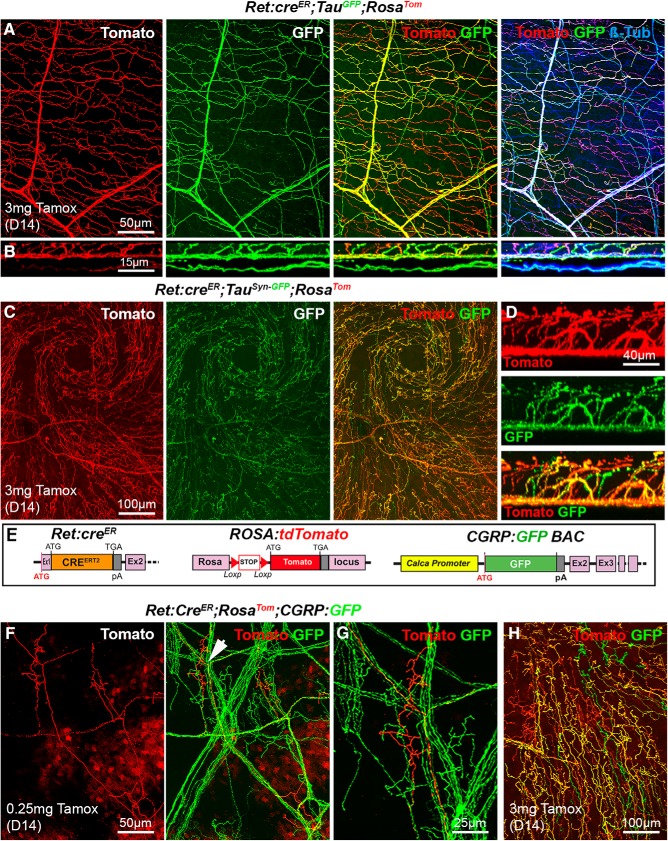
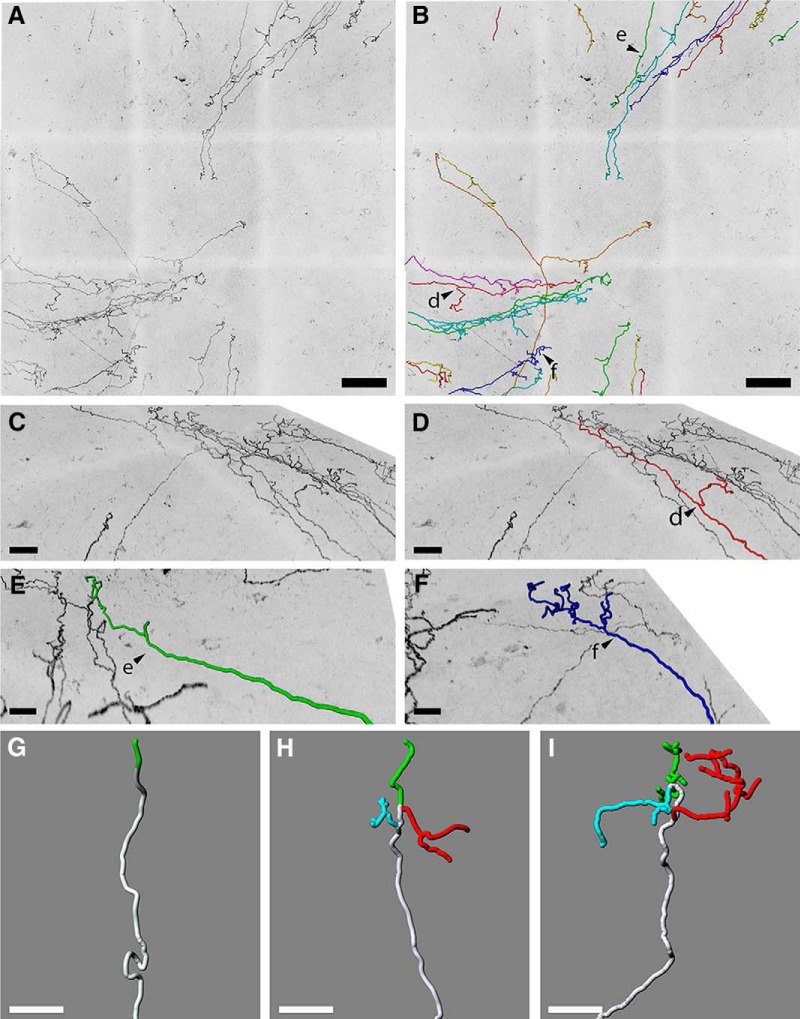
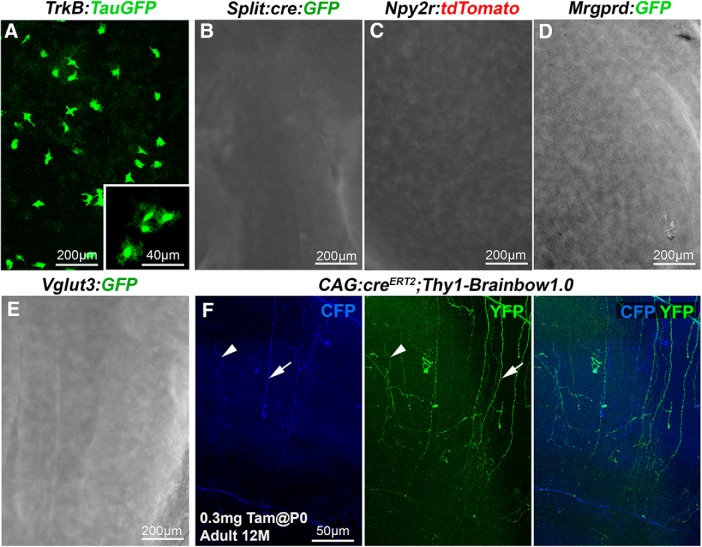
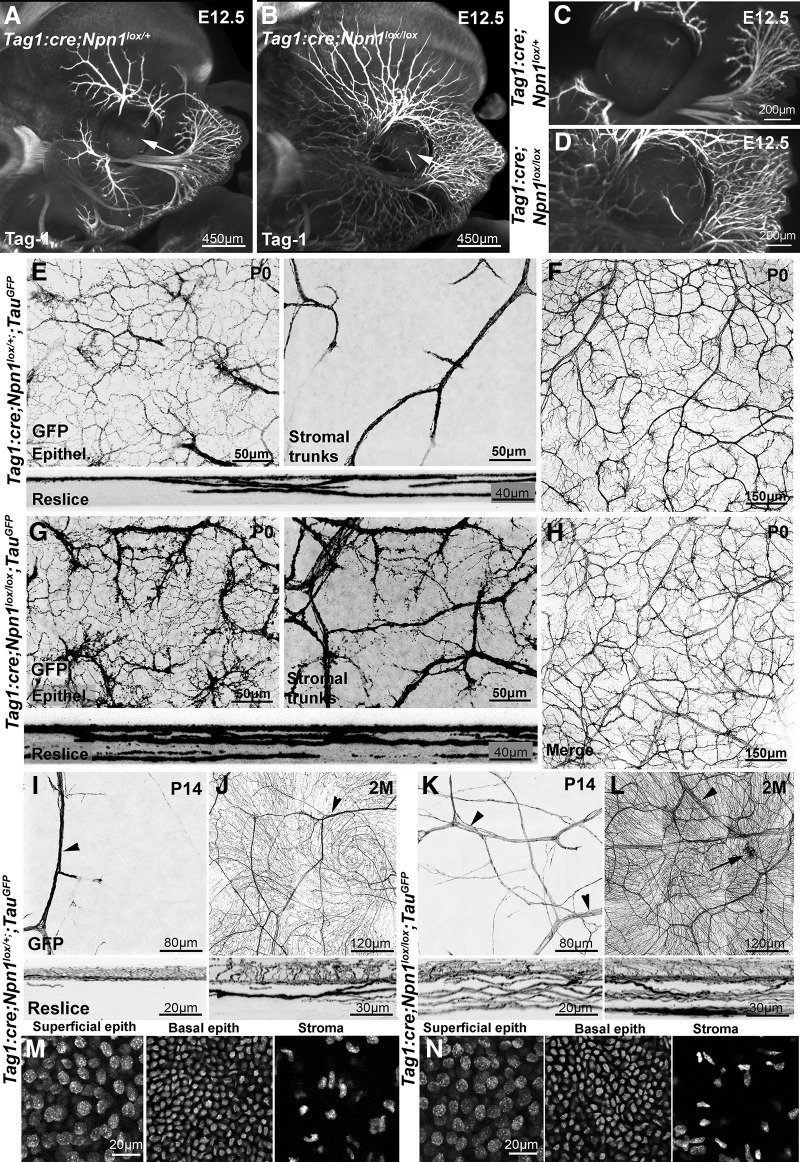
The cornea has the densest sensory innervation of the body, originating primarily from neurons in the trigeminal ganglion. The basic principles of cornea nerve patterning have been established many years ago using classic neuroanatomical methods, such as immunocytochemistry and electrophysiology. Our understanding of the morphology and distribution of the sensory nerves in the skin has considerably progressed over the past few years through the generation and analysis of a variety of genetically modified mouse lines. Surprisingly, these lines were not used to study corneal axons. Here, we have screened a collection of transgenic and knockin mice (of both sexes) to select lines allowing the visualization and genetic manipulation of corneal nerves. We identified multiple lines, including some in which different types of corneal axons can be simultaneously observed with fluorescent proteins expressed in a combinatorial manner. We also provide the first description of the morphology and arborization of single corneal axons and identify three main types of branching pattern. We applied this genetic strategy to the analysis of corneal nerve development and plasticity. We provide direct evidence for a progressive reduction of the density of corneal innervation during aging. We also show that the semaphorin receptor neuropilin-1 acts cell-autonomously to control the development of corneal axons and that early axon guidance defects have long-term consequences on corneal innervation.SIGNIFICANCE STATEMENT We have screened a collection of transgenic and knockin mice and identify lines allowing the visualization and genetic manipulation of corneal nerves. We provide the first description of the arborization pattern of single corneal axons. We also present applications of this genetic strategy to the analysis of corneal nerve development and remodeling during aging.
Keywords: aging; confocal microscopy; cornea; corneal innervation; mouse genetics; neuropilin.
Copyright © 2019 the authors 0270-6474/19/391150-19$15.00/0.
Figures

References
-
- Alcalde I, Íñigo-Portugués A, González-González O, Almaraz L, Artime E, Morenilla-Palao C, Gallar J, Viana F, Merayo-Lloves J, Belmonte C (2018) Morphological and functional changes in TRPM8-expressing corneal cold thermoreceptor neurons during aging and their impact on tearing in mice. J Comp Neurol 520:633–655. 10.1002/cne.24454 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous