

O-GlcNAcylation regulates integrin-mediated cell adhesion and migration via formation of focal adhesion complexes
- PMID: 30587575
- PMCID: PMC6398131
- DOI: 10.1074/jbc.RA118.005923
O-GlcNAcylation regulates integrin-mediated cell adhesion and migration via formation of focal adhesion complexes
Abstract
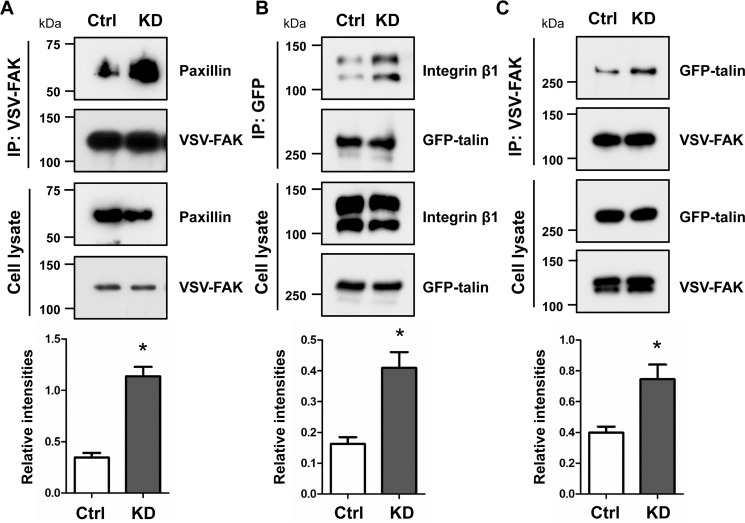
O-GlcNAcylation is a post-translational modification of a protein serine or threonine residue catalyzed by O-GlcNAc transferase (OGT) in the nucleus and cytoplasm. O-GlcNAcylation plays important roles in the cellular signaling that affect the different biological functions of cells, depending upon cell type. However, whether or not O-GlcNAcylation regulates cell adhesion and migration remains unclear. Here, we used the doxycycline-inducible short hairpin RNA (shRNA) system to establish an OGT knockdown (KD) HeLa cell line and found that O-GlcNAcylation is a key regulator for cell adhesion, migration, and focal adhesion (FA) complex formation. The expression levels of OGT and O-GlcNAcylation were remarkably suppressed 24 h after induction of doxycycline. Knockdown of OGT significantly promoted cell adhesion, but it suppressed the cell migration on fibronectin. The immunostaining with paxillin, a marker for FA plaque, clearly showed that the number of FAs was increased in the KD cells compared with that in the control cells. The O-GlcNAcylation levels of paxillin, talin, and focal adhesion kinase were down-regulated in KD cells. Interestingly, the complex formation between integrin β1, focal adhesion kinase, paxillin, and talin was greatly increased in KD cells. Consistently, levels of active integrin β1 were significantly enhanced in KD cells, whereas they were decreased in cells overexpressing OGT. The data suggest a novel regulatory mechanism for O-GlcNAcylation during FA complex formation, which thereby affects integrin activation and integrin-mediated functions such as cell adhesion and migration.
Keywords: O-GlcNAcylation; O-linked N-acetylglucosamine (O-GlcNAc) transferase (OGT); cell adhesion; cell migration; focal adhesions; glycosylation; integrin.
© 2019 Xu et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures






References
-
- Itkonen H. M., Minner S., Guldvik I. J., Sandmann M. J., Tsourlakis M. C., Berge V., Svindland A., Schlomm T., and Mills I. G. (2013) O-GlcNAc transferase integrates metabolic pathways to regulate the stability of c-MYC in human prostate cancer cells. Cancer Res. 73, 5277–5287 10.1158/0008-5472.CAN-13-0549 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
