

Bipolar switching by HCN voltage sensor underlies hyperpolarization activation
- PMID: 30587580
- PMCID: PMC6329955
- DOI: 10.1073/pnas.1816724116
Bipolar switching by HCN voltage sensor underlies hyperpolarization activation
Abstract
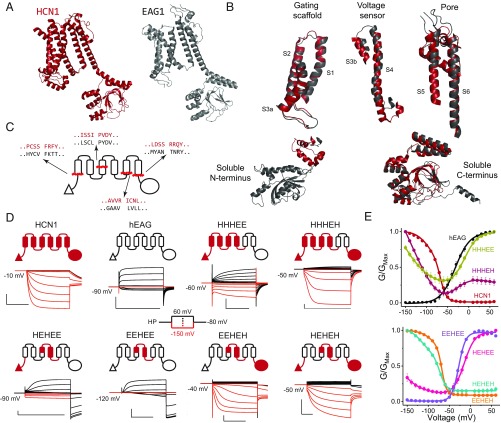
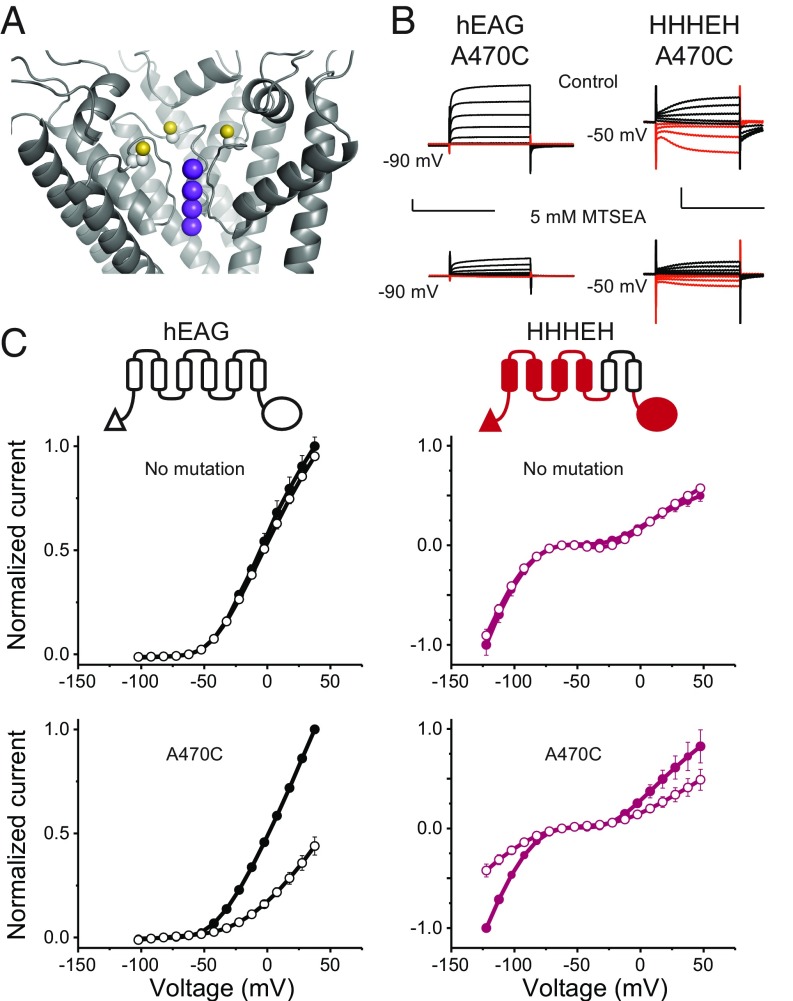
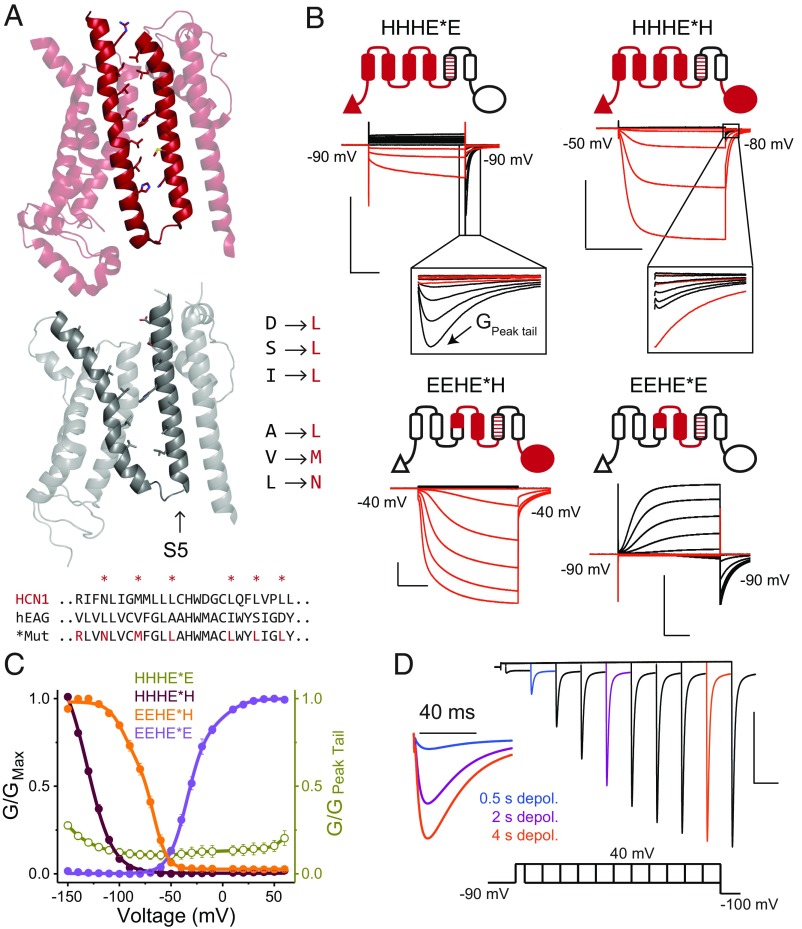
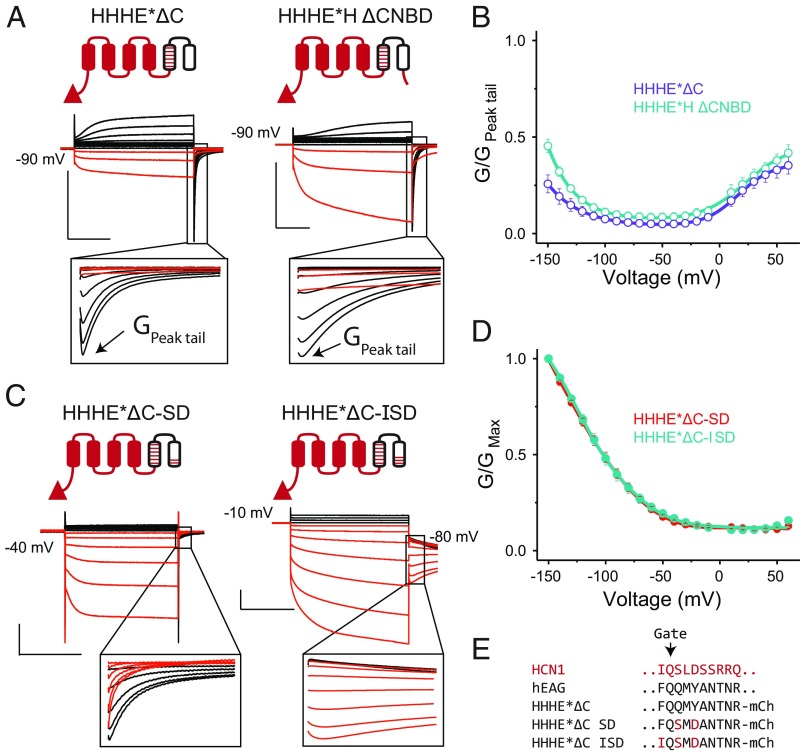
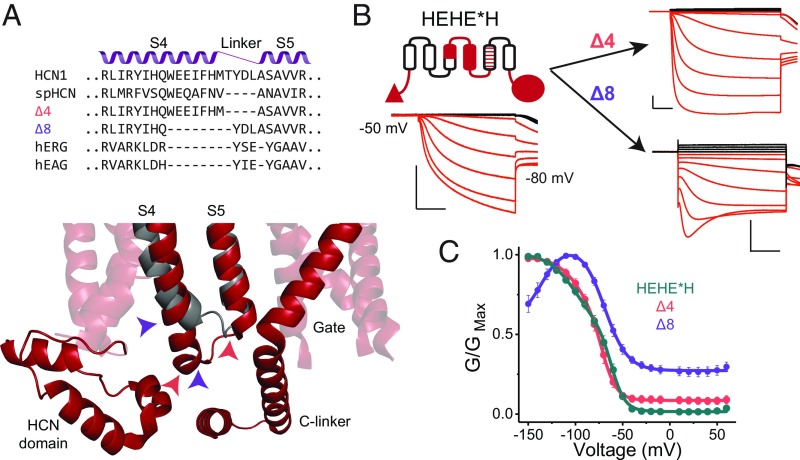
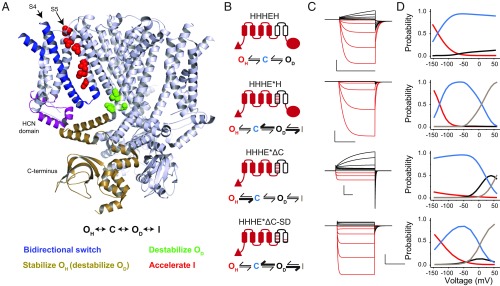
Despite sharing a common architecture with archetypal voltage-gated ion channels (VGICs), hyperpolarization- and cAMP-activated ion (HCN) channels open upon hyperpolarization rather than depolarization. The basic motions of the voltage sensor and pore gates are conserved, implying that these domains are inversely coupled in HCN channels. Using structure-guided protein engineering, we systematically assembled an array of mosaic channels that display the full complement of voltage-activation phenotypes observed in the VGIC superfamily. Our studies reveal that the voltage sensor of the HCN channel has an intrinsic ability to drive pore opening in either direction and that the extra length of the HCN S4 is not the primary determinant for hyperpolarization activation. Tight interactions at the HCN voltage sensor-pore interface drive the channel into an hERG-like inactivated state, thereby obscuring its opening upon depolarization. This structural element in synergy with the HCN cyclic nucleotide-binding domain and specific interactions near the pore gate biases the channel toward hyperpolarization-dependent opening. Our findings reveal an unexpected common principle underpinning voltage gating in the VGIC superfamily and identify the essential determinants of gating polarity.
Keywords: EAG; HCN; depolarization; hyperpolarization; ion channel.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The HCN domain is required for HCN channel cell-surface expression and couples voltage- and cAMP-dependent gating mechanisms.J Biol Chem. 2020 Jun 12;295(24):8164-8173. doi: 10.1074/jbc.RA120.013281. Epub 2020 Apr 27. J Biol Chem. 2020. PMID: 32341127 Free PMC article.
-
Similar voltage-sensor movement in spHCN channels can cause closing, opening, or inactivation.J Gen Physiol. 2023 May 1;155(5):e202213170. doi: 10.1085/jgp.202213170. Epub 2023 Feb 8. J Gen Physiol. 2023. PMID: 36752823 Free PMC article.
-
The S4-S5 linker couples voltage sensing and activation of pacemaker channels.Proc Natl Acad Sci U S A. 2001 Sep 25;98(20):11277-82. doi: 10.1073/pnas.201250598. Epub 2001 Sep 11. Proc Natl Acad Sci U S A. 2001. PMID: 11553787 Free PMC article.
-
Structural insights into the mechanisms of CNBD channel function.J Gen Physiol. 2018 Feb 5;150(2):225-244. doi: 10.1085/jgp.201711898. Epub 2017 Dec 12. J Gen Physiol. 2018. PMID: 29233886 Free PMC article. Review.
-
The structure of the apo cAMP-binding domain of HCN4 - a stepping stone toward understanding the cAMP-dependent modulation of the hyperpolarization-activated cyclic-nucleotide-gated ion channels.FEBS J. 2018 Jun;285(12):2182-2192. doi: 10.1111/febs.14408. Epub 2018 Mar 14. FEBS J. 2018. PMID: 29444387 Review.
Cited by
-
Voltage Sensor Movements during Hyperpolarization in the HCN Channel.Cell. 2019 Dec 12;179(7):1582-1589.e7. doi: 10.1016/j.cell.2019.11.006. Epub 2019 Nov 28. Cell. 2019. PMID: 31787376 Free PMC article.
-
Allosteric signaling in C-linker and cyclic nucleotide-binding domain of HCN2 channels.Biophys J. 2021 Mar 2;120(5):950-963. doi: 10.1016/j.bpj.2021.01.017. Epub 2021 Jan 28. Biophys J. 2021. PMID: 33515603 Free PMC article.
-
The HCN channel voltage sensor undergoes a large downward motion during hyperpolarization.Nat Struct Mol Biol. 2019 Aug;26(8):686-694. doi: 10.1038/s41594-019-0259-1. Epub 2019 Jul 8. Nat Struct Mol Biol. 2019. PMID: 31285608 Free PMC article.
-
Review: HCN Channels in the Heart.Curr Cardiol Rev. 2022;18(4):e040222200836. doi: 10.2174/1573403X18666220204142436. Curr Cardiol Rev. 2022. PMID: 35125083 Free PMC article. Review.
-
hERG Function in Light of Structure.Biophys J. 2020 Feb 25;118(4):790-797. doi: 10.1016/j.bpj.2019.10.010. Epub 2019 Oct 18. Biophys J. 2020. PMID: 31669064 Free PMC article. Review.
References
-
- Bezanilla F. The voltage sensor in voltage-dependent ion channels. Physiol Rev. 2000;80:555–592. - PubMed
-
- Ludwig A, Zong X, Jeglitsch M, Hofmann F, Biel M. A family of hyperpolarization-activated mammalian cation channels. Nature. 1998;393:587–591. - PubMed
-
- Santoro B, et al. Identification of a gene encoding a hyperpolarization-activated pacemaker channel of brain. Cell. 1998;93:717–729. - PubMed
-
- Brown HF, DiFrancesco D, Noble SJ. How does adrenaline accelerate the heart? Nature. 1979;280:235–236. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
