

LT-IIc, A Bacterial Type II Heat-Labile Enterotoxin, Induces Specific Lethality in Triple Negative Breast Cancer Cells by Modulation of Autophagy and Induction of Apoptosis and Necroptosis
- PMID: 30587795
- PMCID: PMC6337683
- DOI: 10.3390/ijms20010085
LT-IIc, A Bacterial Type II Heat-Labile Enterotoxin, Induces Specific Lethality in Triple Negative Breast Cancer Cells by Modulation of Autophagy and Induction of Apoptosis and Necroptosis
Abstract
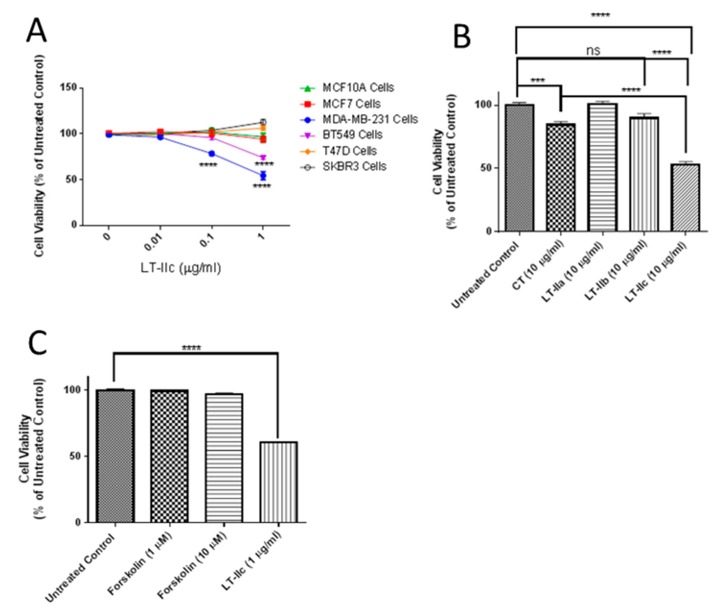
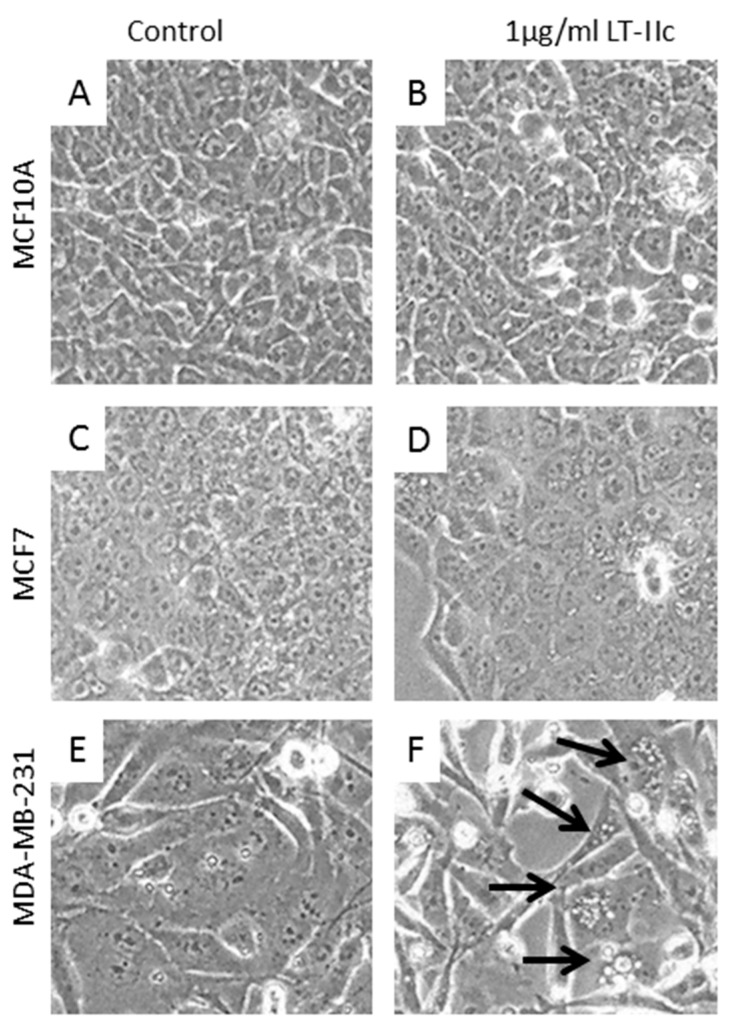
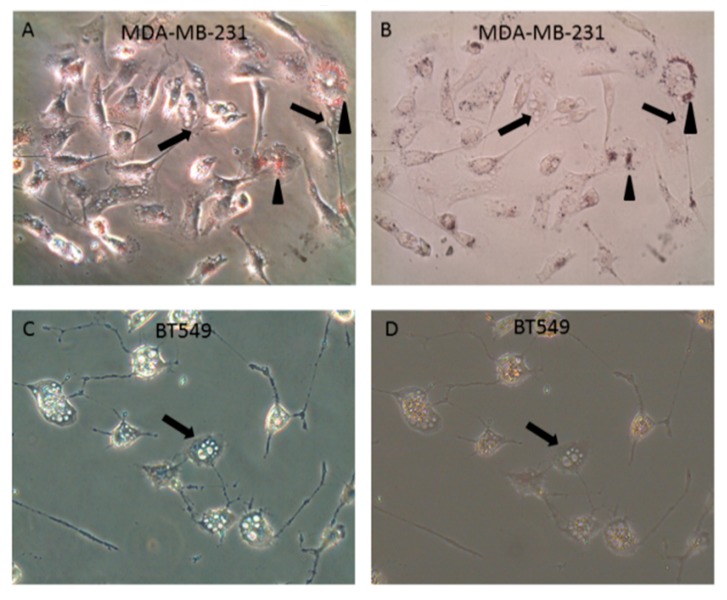
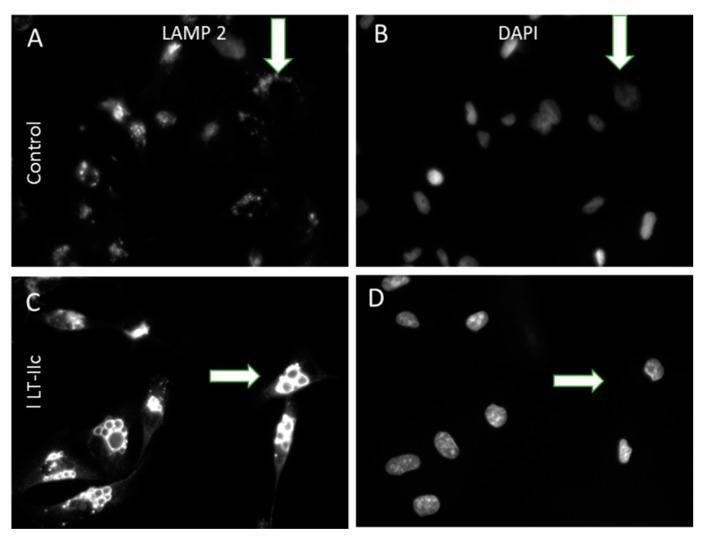
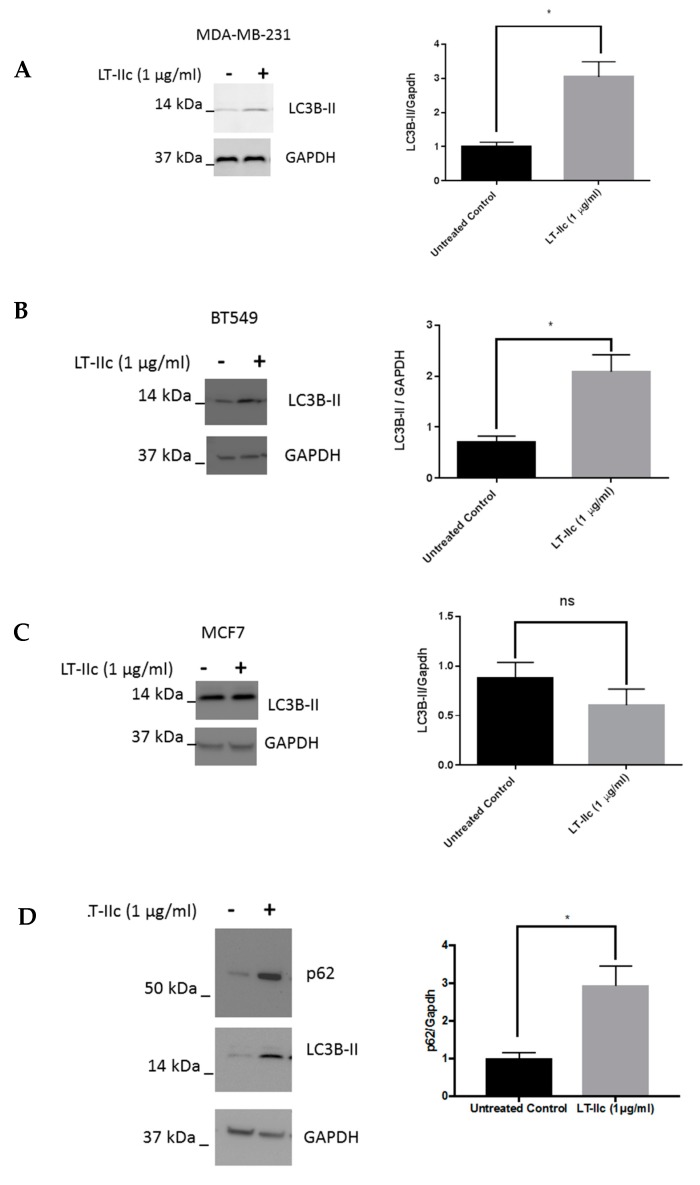
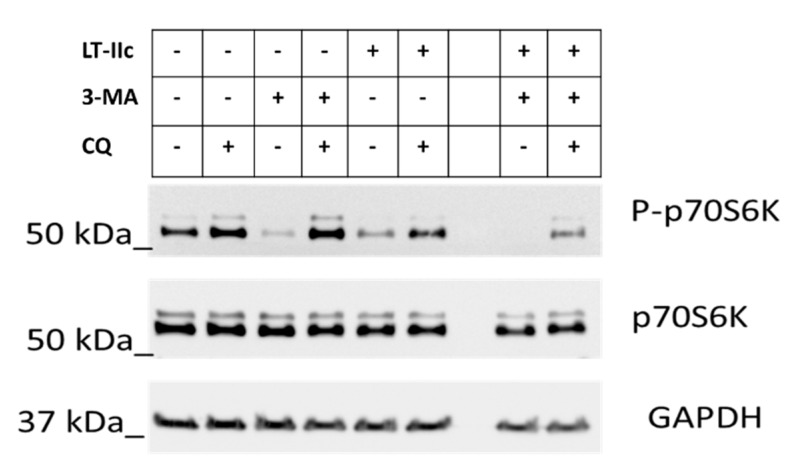
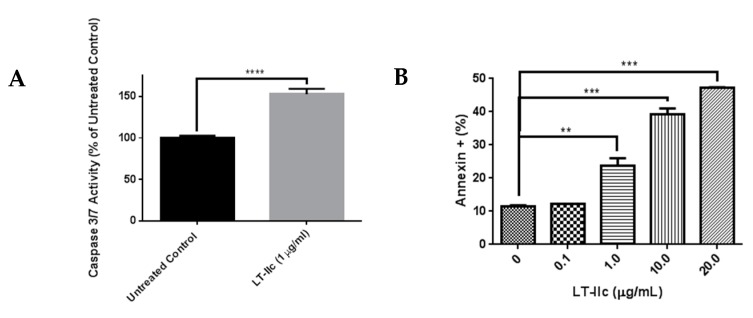
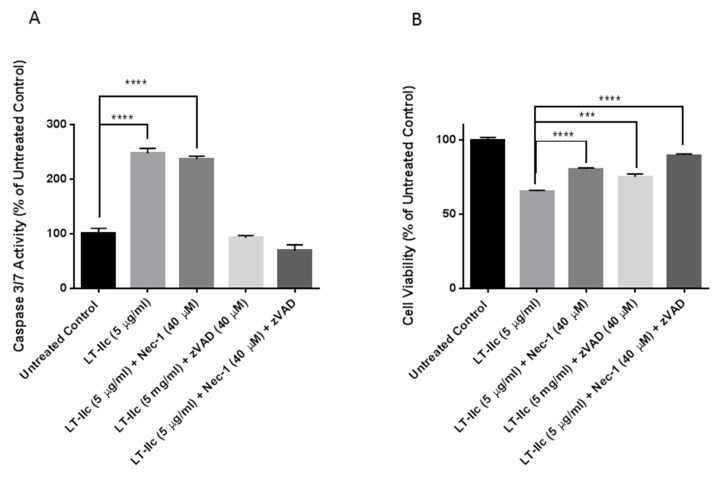
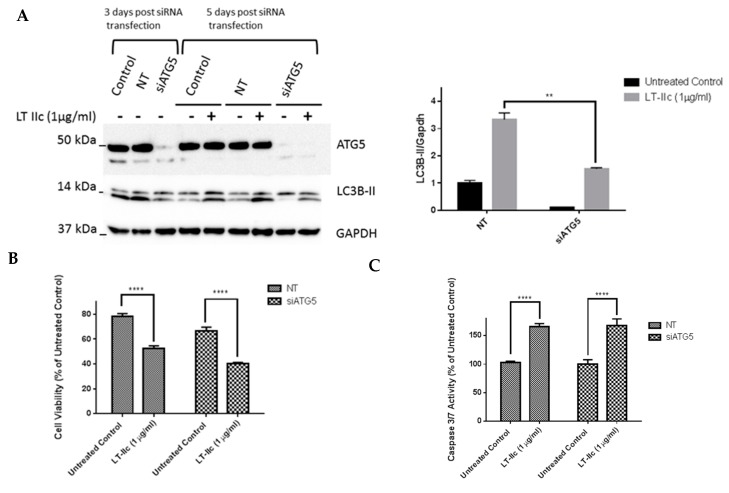
Triple negative breast cancer (TNBC) remains a serious health problem with poor prognosis and limited therapeutic options. To discover novel approaches to treat TNBC, we screened cholera toxin (CT) and the members of the bacterial type II heat-labile enterotoxin family (LT-IIa, LT-IIb, and LT-IIc) for cytotoxicity in TNBC cells. Only LT-IIc significantly reduced viability of the TNBC cell lines BT549 and MDA-MB-231 (IC50 = 82.32 nM). LT-IIc had no significant cytotoxic effect on MCF10A (IC50 = 2600 nM), a non-tumorigenic breast epithelial cell line, and minimal effects on MCF7 and T47D, ER⁺ cells, or SKBR-3 cells, HER2⁺ cells. LT-IIc stimulated autophagy through inhibition of the mTOR pathway, while simultaneously inhibiting autophagic progression, as seen by accumulation of LC3B-II and p62. Morphologically, LT-IIc induced the formation of enlarged LAMP2+ autolysosomes, which was blocked by co-treatment with bafilomycin A1. LT-IIc induced apoptosis as demonstrated by the increase in caspase 3/7 activity and Annexin V staining. Co-treatment with necrostatin-1, however, demonstrated that the lethal response of LT-IIc is elicited, in part, by concomitant induction of necroptosis. Knockdown of ATG-5 failed to rescue LT-IIc-induced cytotoxicity, suggesting LT-IIc can exert its cytotoxic effects downstream or independently of autophagophore initiation. Collectively, these experiments demonstrate that LT-IIc acts bifunctionally, inducing autophagy, while simultaneously blocking autolysosomal progression in TNBC cells, inducing a specific cytotoxicity in this breast cancer subtype.
Keywords: LT-IIc; apoptosis; autophagy; bacterial enterotoxins; breast cancer; heat-labile enterotoxins; necroptosis; triple-negative breast cancer.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Havas H.F., Donnelly A.J. Mixed bacterial toxins in the treatment of tumors. IV. Response of methylcholanthrene-induced, spontaneous, and transplanted tumors in mice. Cancer Res. 1961;21:17–25. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
