

Structure and two-metal mechanism of fungal tRNA ligase
- PMID: 30590734
- PMCID: PMC6379707
- DOI: 10.1093/nar/gky1275
Structure and two-metal mechanism of fungal tRNA ligase
Abstract
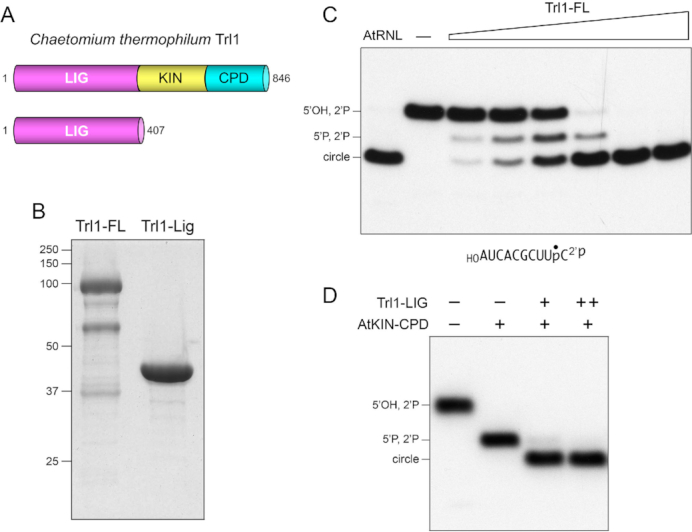
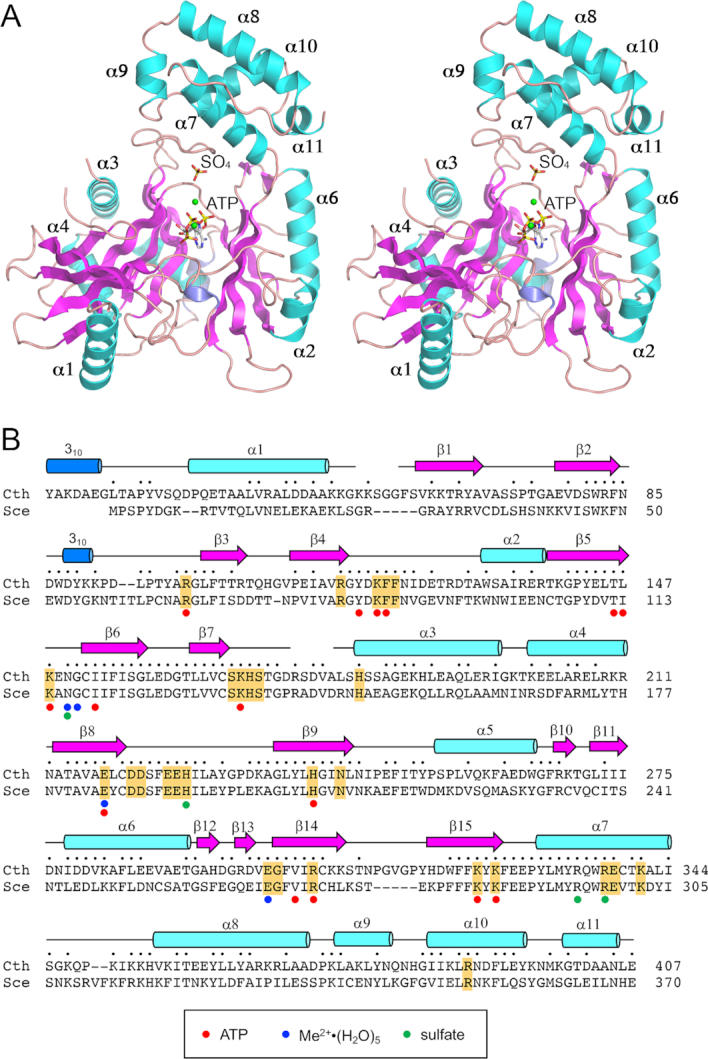
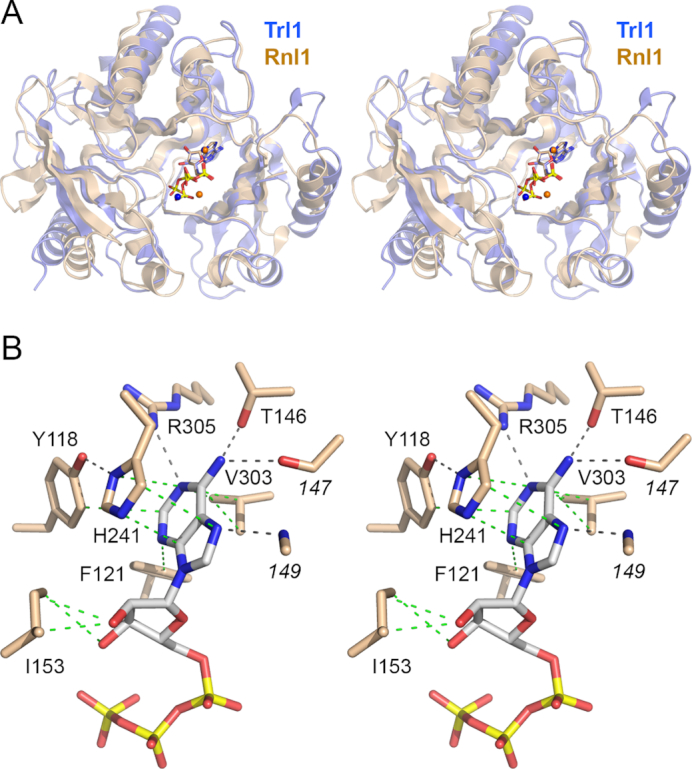
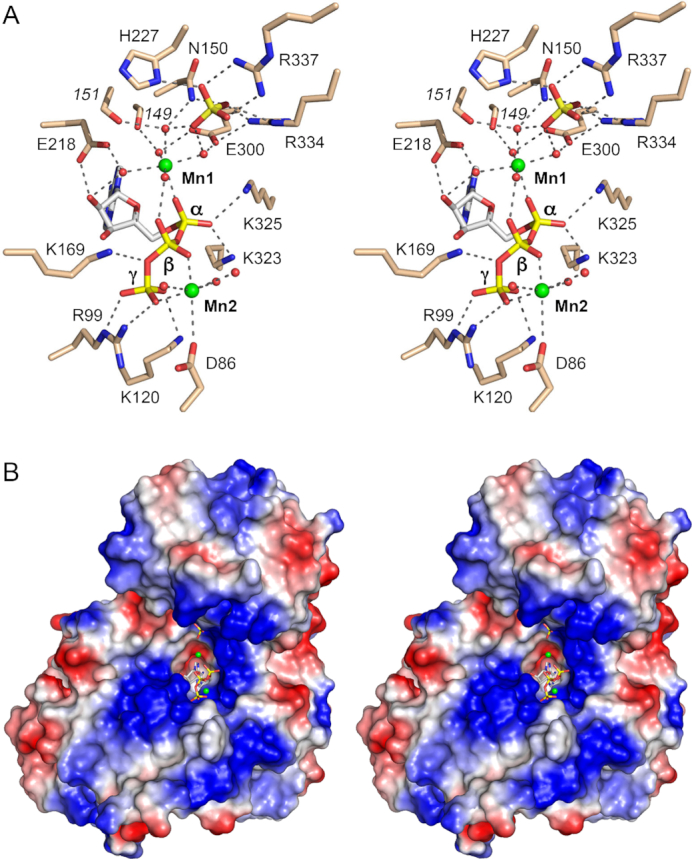
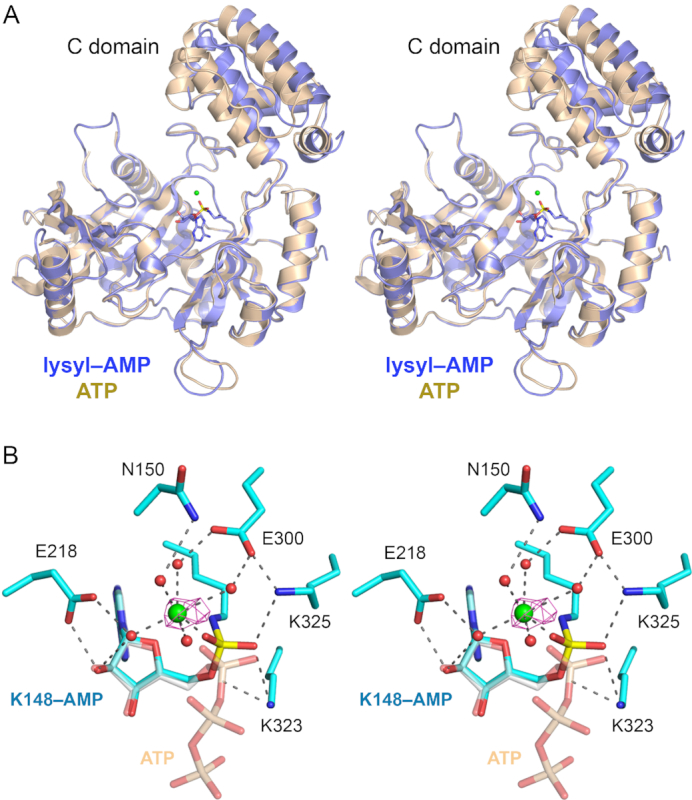
Fungal tRNA ligase (Trl1) is an essential enzyme that repairs RNA breaks with 2',3'-cyclic-PO4 and 5'-OH ends inflicted during tRNA splicing and non-canonical mRNA splicing in the fungal unfolded protein response. Trl1 is composed of C-terminal cyclic phosphodiesterase (CPD) and central GTP-dependent polynucleotide kinase (KIN) domains that heal the broken ends to generate the 3'-OH,2'-PO4 and 5'-PO4 termini required for sealing by an N-terminal ATP-dependent ligase domain (LIG). Here we report crystal structures of the Trl1-LIG domain from Chaetomium thermophilum at two discrete steps along the reaction pathway: the covalent LIG-(lysyl-Nζ)-AMP•Mn2+ intermediate and a LIG•ATP•(Mn2+)2 Michaelis complex. The structures highlight a two-metal mechanism whereby a penta-hydrated metal complex stabilizes the transition state of the ATP α phosphate and a second metal bridges the β and γ phosphates to help orient the pyrophosphate leaving group. A LIG-bound sulfate anion is a plausible mimetic of the essential RNA terminal 2'-PO4. Trl1-LIG has a distinctive C-terminal domain that instates fungal Trl1 as the founder of an Rnl6 clade of ATP-dependent RNA ligase. We discuss how the Trl1-LIG structure rationalizes the large body of in vivo structure-function data for Saccharomyces cerevisiae Trl1.
© The Author(s) 2018. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Abelson J., Trotta C.R., Li H.. tRNA splicing. J. Biol. Chem. 1998; 273:12685–12688. - PubMed
-
- Sidrauski C., Cox J.S., Walter P.. tRNA ligase is required for regulated mRNA splicing in the unfolded protein response. Cell. 1996; 87:405–413. - PubMed
-
- Greer C.L., Peebles C.L., Gegenheimer P., Abelson J.. Mechanism of action of a yeast RNA ligase in tRNA splicing. Cell. 1983; 32:537–546. - PubMed
-
- Apostol B.L., Westaway S.K., Abelson J., Greer C.L.. Deletion analysis of a multifunctional yeast tRNA ligase polypeptide: identification of essential and dispensable functional domains. J. Biol. Chem. 1991; 266:7445–7455. - PubMed
-
- Sawaya R., Schwer B., Shuman S.. Genetic and biochemical analysis of the functional domains of yeast tRNA ligase. J. Biol. Chem. 2003; 278:43298–43398. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
