

Resolution of the Holliday junction recombination intermediate by human GEN1 at the single-molecule level
- PMID: 30590761
- PMCID: PMC6393306
- DOI: 10.1093/nar/gky1280
Resolution of the Holliday junction recombination intermediate by human GEN1 at the single-molecule level
Abstract
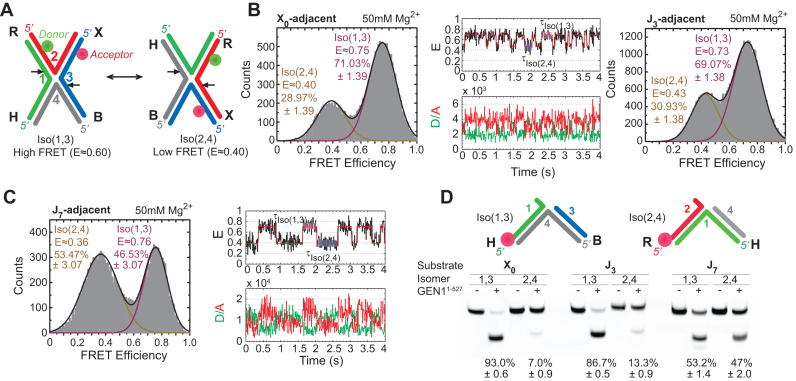
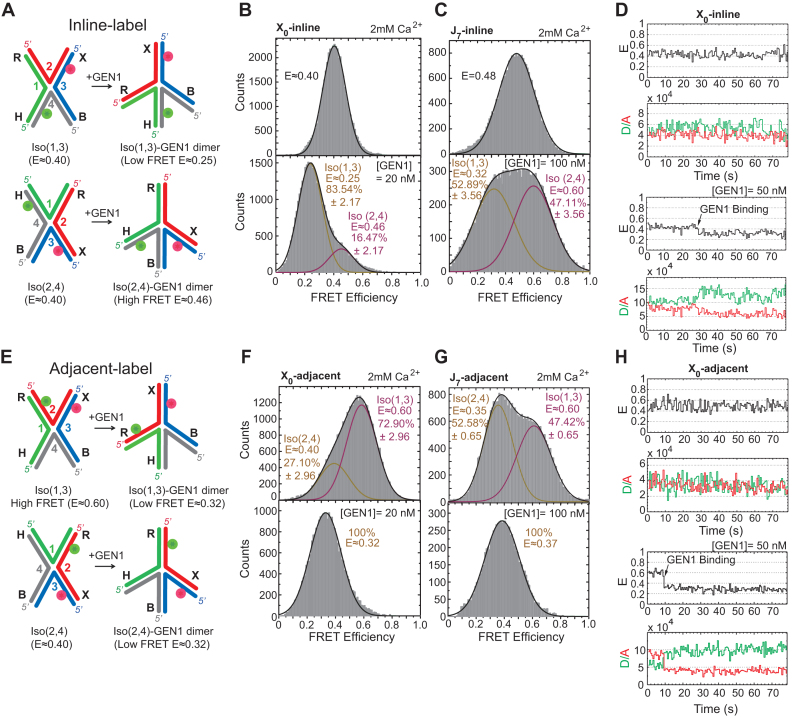
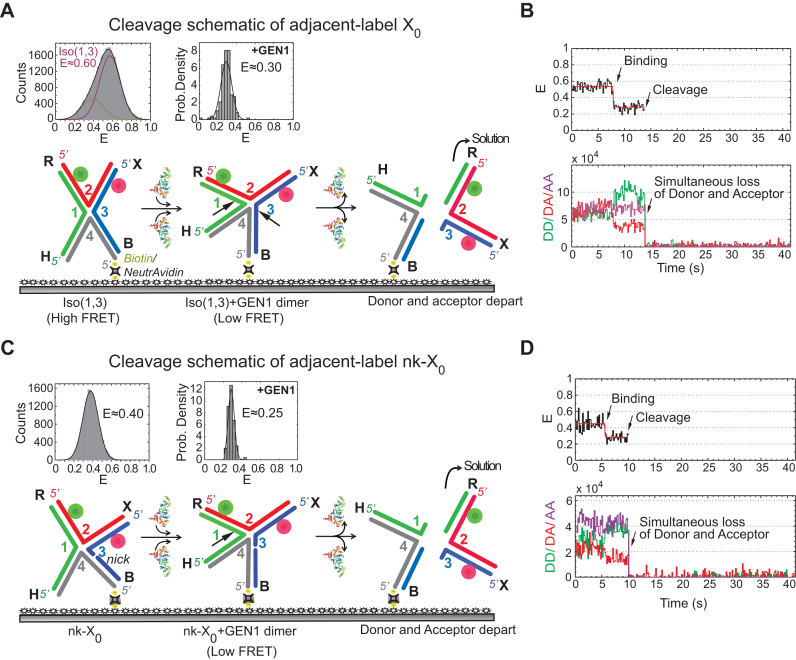
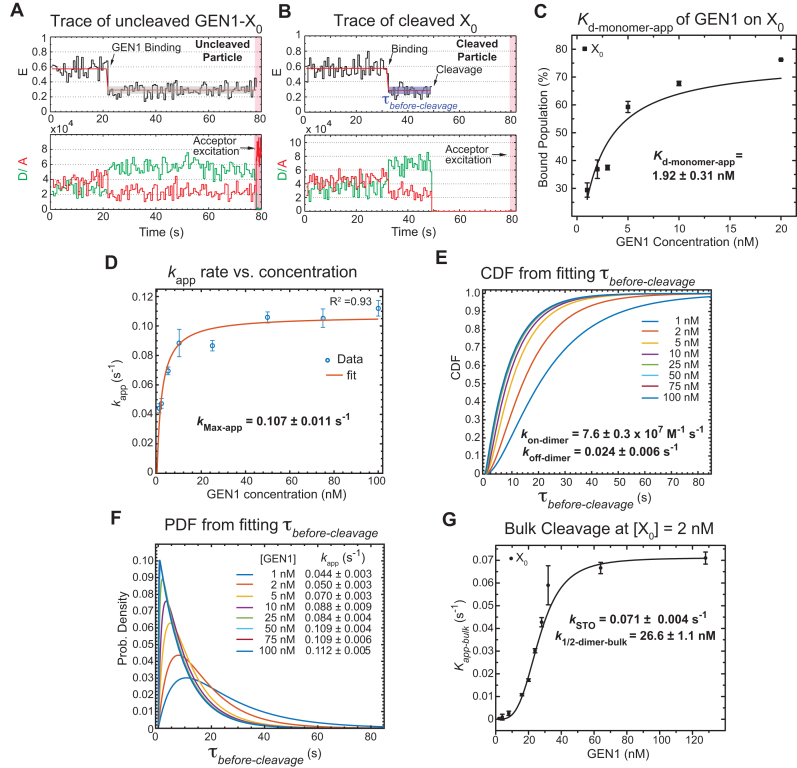
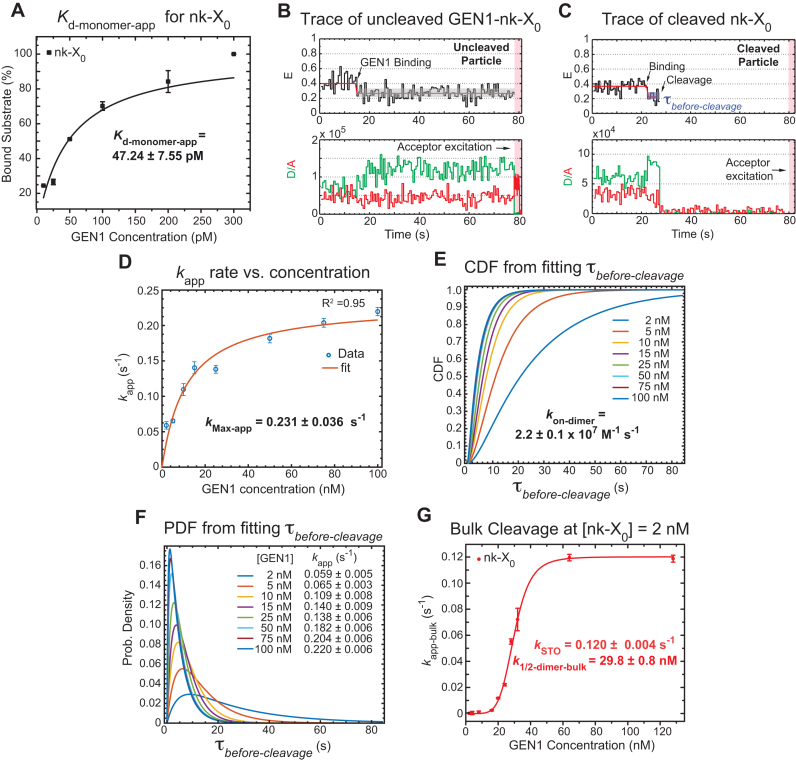
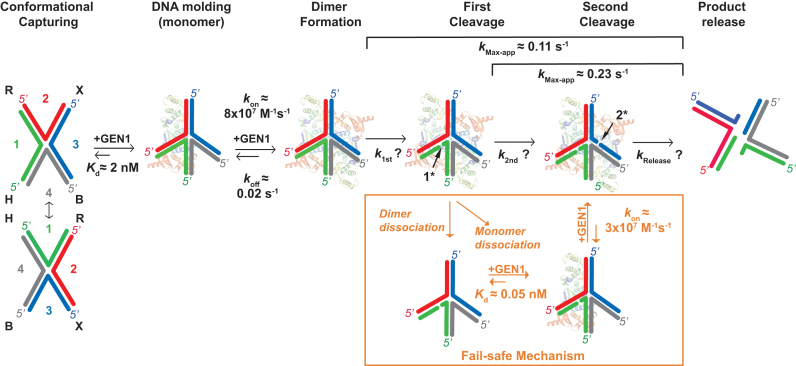
Human GEN1 is a cytosolic homologous recombination protein that resolves persisting four-way Holliday junctions (HJ) after the dissolution of the nuclear membrane. GEN1 dimerization has been suggested to play key role in the resolution of the HJ, but the kinetic details of its reaction remained elusive. Here, single-molecule FRET shows how human GEN1 binds the HJ and always ensures its resolution within the lifetime of the GEN1-HJ complex. GEN1 monomer generally follows the isomer bias of the HJ in its initial binding and subsequently distorts it for catalysis. GEN1 monomer remains tightly bound with no apparent dissociation until GEN1 dimer is formed and the HJ is fully resolved. Fast on- and slow off-rates of GEN1 dimer and its increased affinity to the singly-cleaved HJ enforce the forward reaction. Furthermore, GEN1 monomer binds singly-cleaved HJ tighter than intact HJ providing a fail-safe mechanism if GEN1 dimer or one of its monomers dissociates after the first cleavage. The tight binding of GEN1 monomer to intact- and singly-cleaved HJ empowers it as the last resort to process HJs that escape the primary mechanisms.
© The Author(s) 2018. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Holliday R. Mechanism for gene conversion in fungi. Genet. Res. 1964; 5:282. - PubMed
-
- West S.C. Formation, translocationand resolution of Holliday junctions during homologous genetic recombination. Philos. Trans.R. Soc. Lond., B, Biol. Sci. 1995; 347:21. - PubMed
-
- Cox M.M., Goodman M.F., Kreuzer K.N., Sherratt D.J., Sandler S.J., Marians K.J.. The importance of repairing stalled replication forks. Nature. 2000; 404:37. - PubMed
-
- West S.C. Molecular views of recombination proteins and their control. Nat. Rev. Mol. Cell Biol. 2003; 4:435. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
