

Insights into the Bidirectional Properties of the Sheep-Deer Prion Transmission Barrier
- PMID: 30592012
- PMCID: PMC6614146
- DOI: 10.1007/s12035-018-1443-8
Insights into the Bidirectional Properties of the Sheep-Deer Prion Transmission Barrier
Abstract
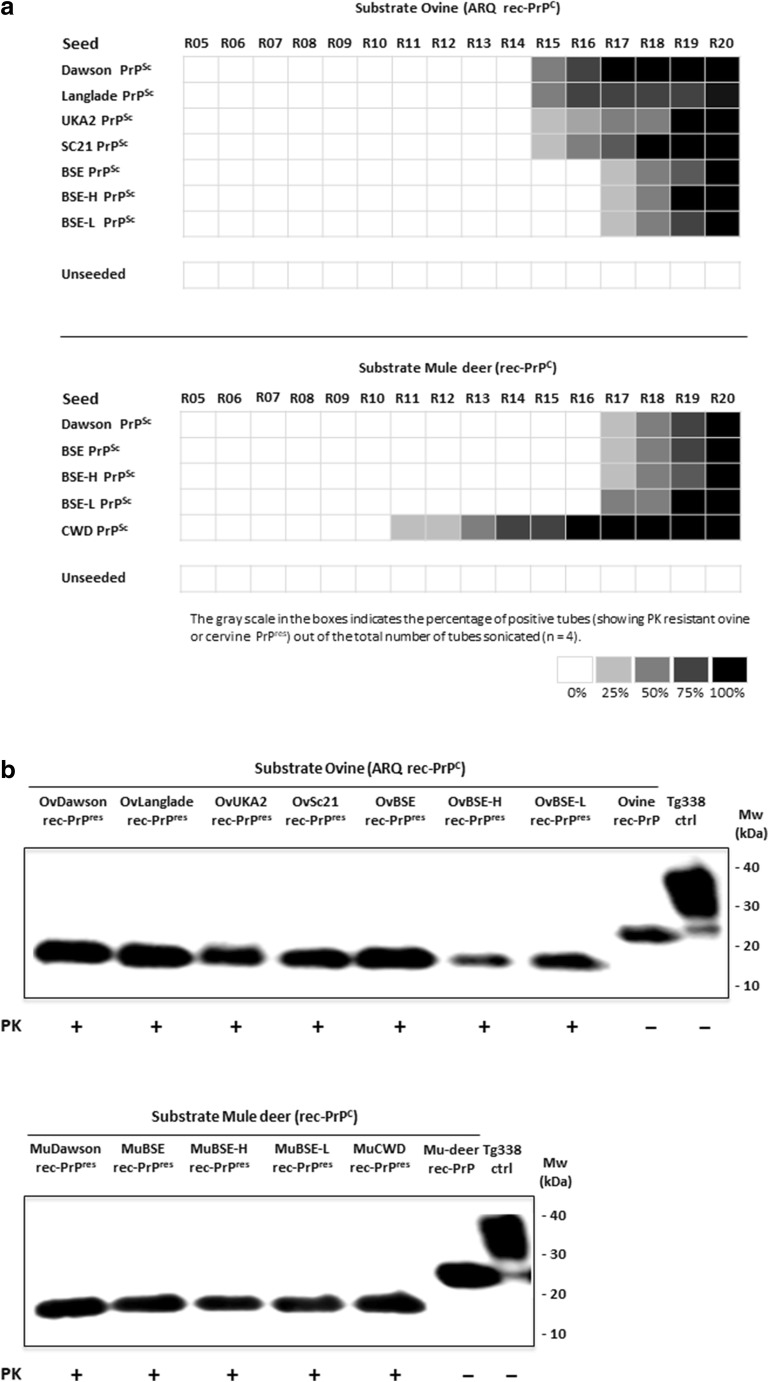
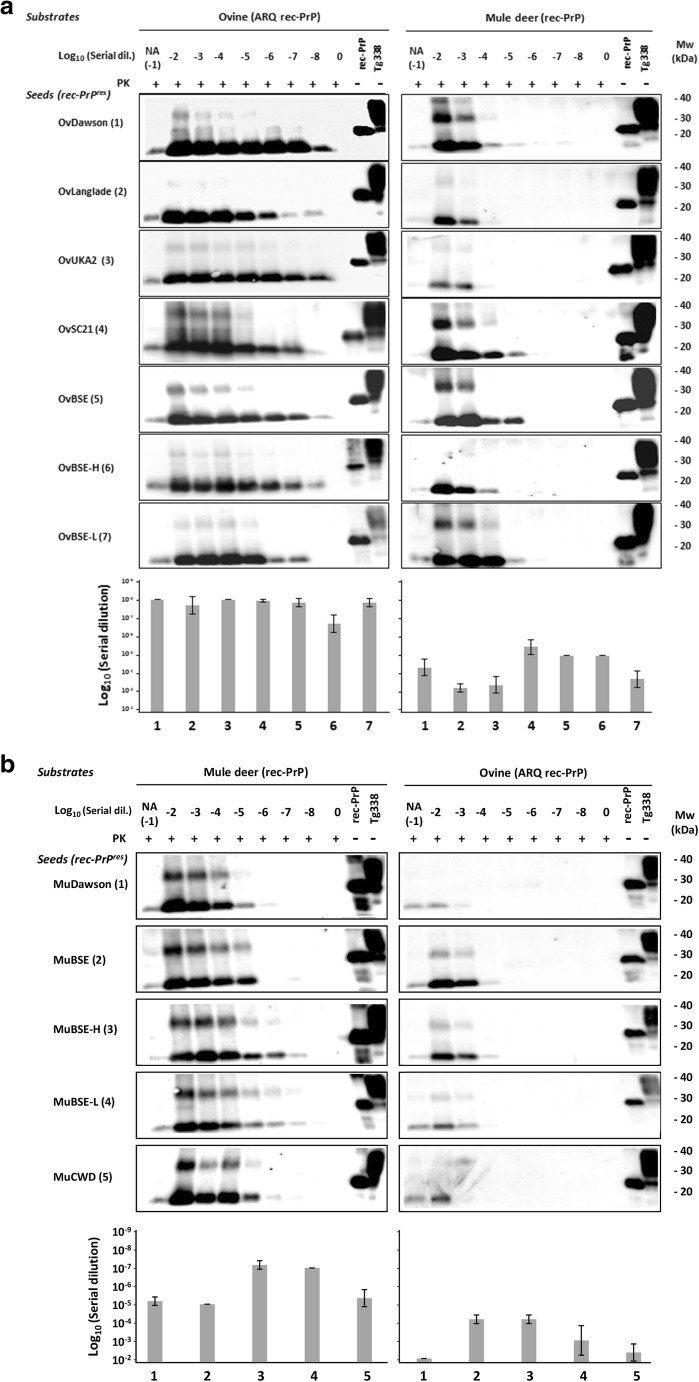
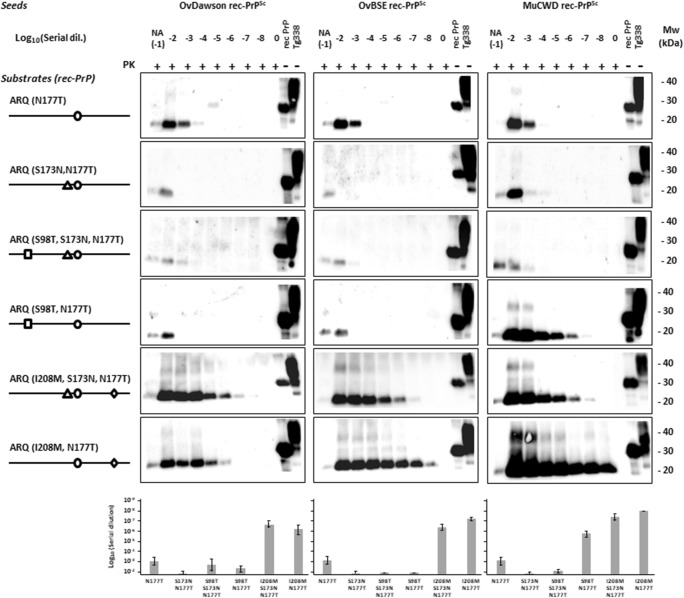
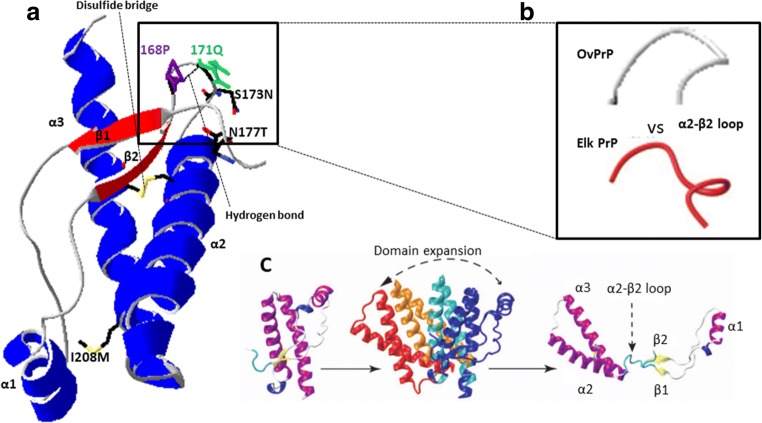
The large chronic wasting disease (CWD)-affected cervid population in the USA and Canada, and the risk of the disease being transmitted to humans through intermediate species, is a highly worrying issue that is still poorly understood. In this case, recombinant protein misfolding cyclic amplification was used to determine, in vitro, the relevance of each individual amino acid on cross-species prion transmission. Others and we have found that the β2-α2 loop is a key modulator of transmission barriers between species and markedly influences infection by sheep scrapie, bovine spongiform encephalopathy (BSE), or elk CWD. Amino acids that differentiate ovine and deer normal host prion protein (PrPC) and associated with structural rigidity of the loop β2-α2 (S173N, N177T) appear to confer resistance to some prion diseases. However, addition of methionine at codon 208 together with the previously described rigid loop substitutions seems to hide a key in this species barrier, as it makes sheep recombinant prion protein highly susceptible to CWD-induced misfolding. These studies indicate that interspecies prion transmission is not only governed just by the β2-α2 loop amino acid sequence but also by its interactions with the α3-helix as shown by substitution I208M. Transmissible spongiform encephalopathies, characterized by long incubation periods and spongiform changes associated with neuronal loss in the brain, have been described in several mammalian species appearing either naturally (scrapie in sheep and goats, bovine spongiform encephalopathy in cattle, chronic wasting disease in cervids, Creutzfeldt-Jakob disease in humans) or by experimental transmission studies (scrapie in mice and hamsters). Much of the pathogenesis of the prion diseases has been determined in the last 40 years, such as the etiological agent or the fact that prions occur as different strains that show distinct biological and physicochemical properties. However, there are many unanswered questions regarding the strain phenomenon and interspecies transmissibility. To assess the risk of interspecies transmission between scrapie and chronic wasting disease, an in vitro prion propagation method has been used. This technique allows to predict the amino acids preventing the transmission between sheep and deer prion diseases.
Keywords: Chronic wasting disease (CWD); In vitro propagation; PMCA; Prion diseases; Scrapie; Transmissible spongiform encephalopathy (TSE); Transmission barrier.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
In Vitro Approach To Identify Key Amino Acids in Low Susceptibility of Rabbit Prion Protein to Misfolding.J Virol. 2017 Nov 30;91(24):e01543-17. doi: 10.1128/JVI.01543-17. Print 2017 Dec 15. J Virol. 2017. PMID: 28978705 Free PMC article.
-
Experimental interspecies transmission studies of the transmissible spongiform encephalopathies to cattle: comparison to bovine spongiform encephalopathy in cattle.J Vet Diagn Invest. 2011 May;23(3):407-20. doi: 10.1177/1040638711403404. J Vet Diagn Invest. 2011. PMID: 21908269 Review.
-
Prion transmission prevented by modifying the β2-α2 loop structure of host PrPC.J Neurosci. 2014 Jan 15;34(3):1022-7. doi: 10.1523/JNEUROSCI.4636-13.2014. J Neurosci. 2014. PMID: 24431459 Free PMC article.
-
Infectious prions in brains and muscles of domestic pigs experimentally challenged with the BSE, scrapie, and CWD agents.mBio. 2025 Aug 18:e0180025. doi: 10.1128/mbio.01800-25. Online ahead of print. mBio. 2025. PMID: 40823831
-
Chronic wasting disease of cervids.Curr Top Microbiol Immunol. 2004;284:193-214. doi: 10.1007/978-3-662-08441-0_8. Curr Top Microbiol Immunol. 2004. PMID: 15148993 Review.
Cited by
-
Understanding Intra-Species and Inter-Species Prion Conversion and Zoonotic Potential Using Protein Misfolding Cyclic Amplification.Front Aging Neurosci. 2021 Aug 3;13:716452. doi: 10.3389/fnagi.2021.716452. eCollection 2021. Front Aging Neurosci. 2021. PMID: 34413769 Free PMC article. Review.
-
Overview of North American Isolates of Chronic Wasting Disease Used for Strain Research.Pathogens. 2025 Mar 4;14(3):250. doi: 10.3390/pathogens14030250. Pathogens. 2025. PMID: 40137736 Free PMC article. Review.
-
Polymorphism of prion protein gene (PRNP) in Nigerian sheep.Prion. 2023 Dec;17(1):44-54. doi: 10.1080/19336896.2023.2186767. Prion. 2023. PMID: 36892181 Free PMC article.
-
Involvement of N- and C-terminal region of recombinant cervid prion protein in its reactivity to CWD and atypical BSE prions in real-time quaking-induced conversion reaction in the presence of high concentrations of tissue homogenates.Prion. 2020 Dec;14(1):283-295. doi: 10.1080/19336896.2020.1858694. Prion. 2020. PMID: 33345717 Free PMC article.
-
Understanding the key features of the spontaneous formation of bona fide prions through a novel methodology that enables their swift and consistent generation.Acta Neuropathol Commun. 2023 Sep 7;11(1):145. doi: 10.1186/s40478-023-01640-8. Acta Neuropathol Commun. 2023. PMID: 37679832 Free PMC article.
References
-
- Prusiner SB. Novel proteinaceous infectious particles cause scrapie. Science. 1982;216:136–144. - PubMed
-
- Detwiler LA, Baylis M. The epidemiology of scrapie. Rev Sci Tech. 2003;22:121–143. - PubMed
-
- Hunter AJ, Caulfield MP, Kimberlin RH. Learning ability of mice infected with different strains of scrapie. Physiol Behav. 1986;36:1089–1092. - PubMed
-
- Bessen RA, Kocisko DA, Raymond GJ, Nandan S, Lansbury PT, Caughey B. Non-genetic propagation of strain-specific properties of scrapie prion protein. Nature. 1995;375:698–700. - PubMed
-
- Williams ES, Young S. Chronic wasting disease of captive mule deer: a spongiform encephalopathy. J Wildl Dis. 1980;16:89–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
