

Evolutionary, structural and functional analysis of the caleosin/peroxygenase gene family in the Fungi
- PMID: 30593269
- PMCID: PMC6309107
- DOI: 10.1186/s12864-018-5334-1
Evolutionary, structural and functional analysis of the caleosin/peroxygenase gene family in the Fungi
Abstract
Background: Caleosin/peroxygenases, CLO/PXG, (designated PF05042 in Pfam) are a group of genes/proteins with anomalous distributions in eukaryotic taxa. We have previously characterised CLO/PXGs in the Viridiplantae. The aim of this study was to investigate the evolution and functions of the CLO/PXGs in the Fungi and other non-plant clades and to elucidate the overall origin of this gene family.
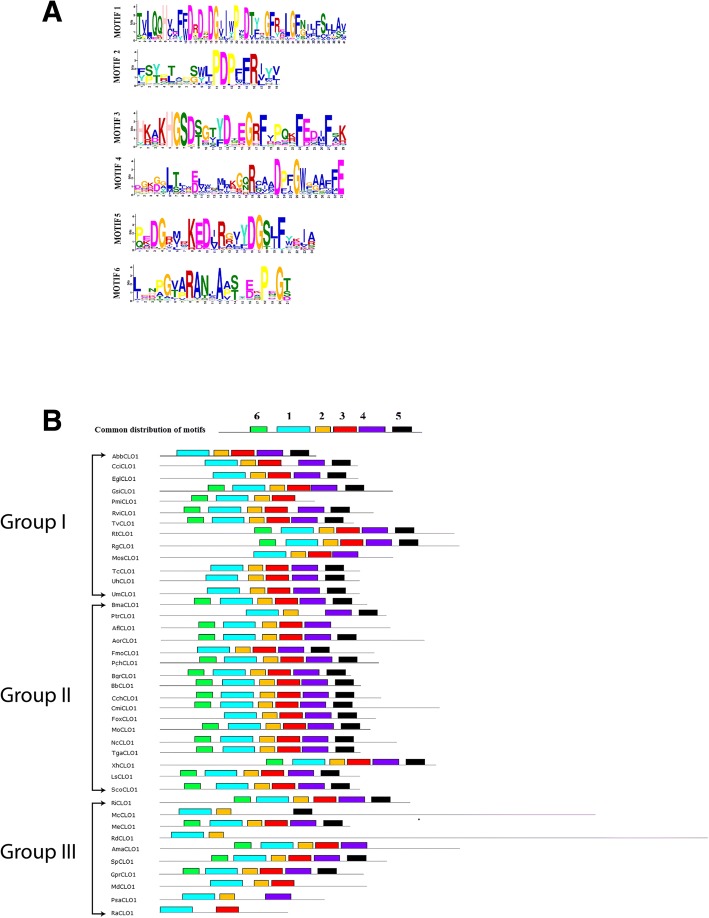
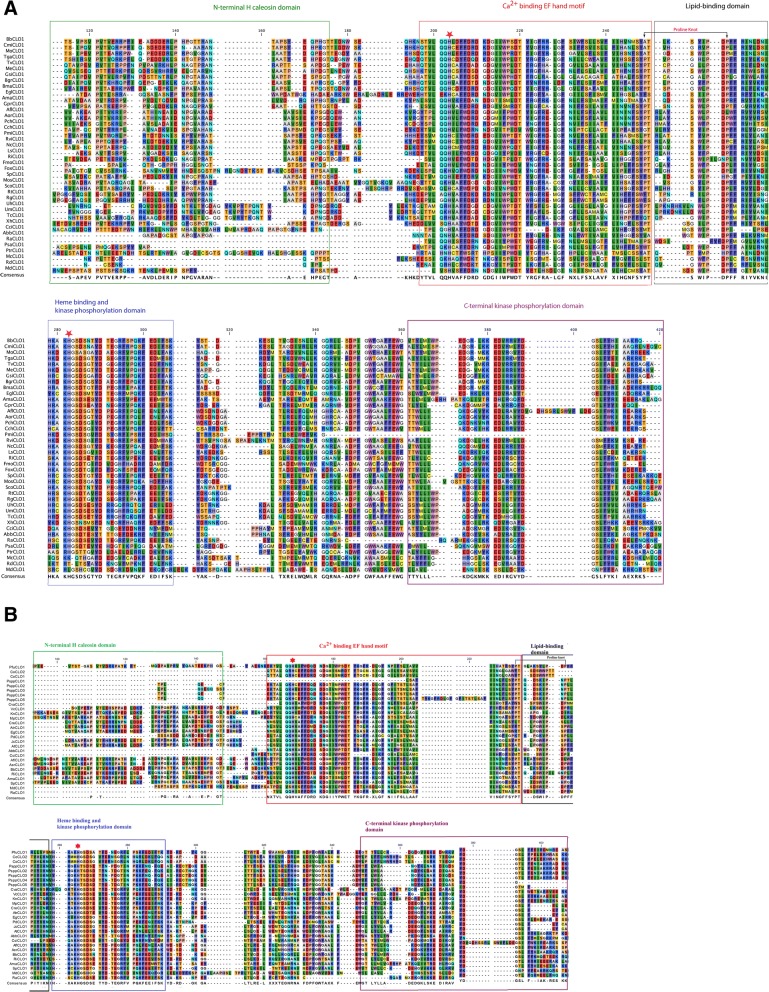
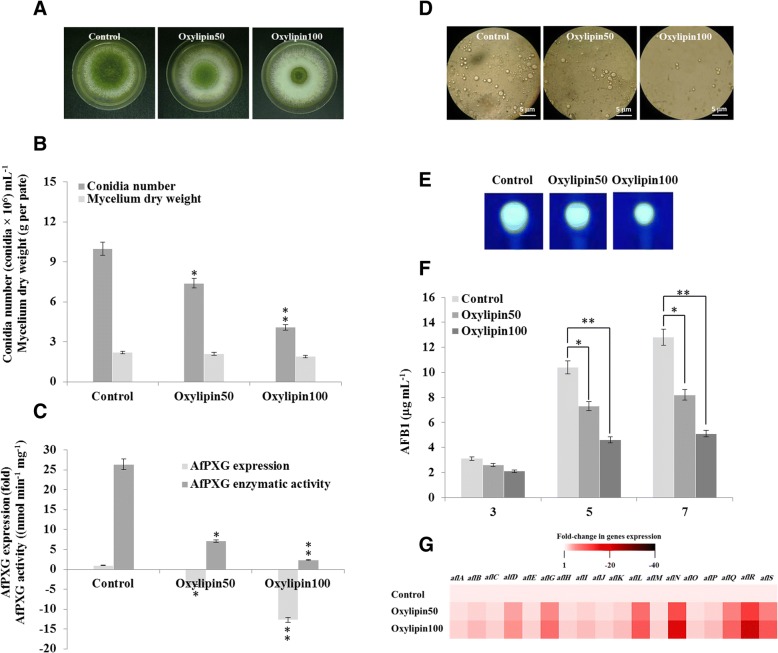
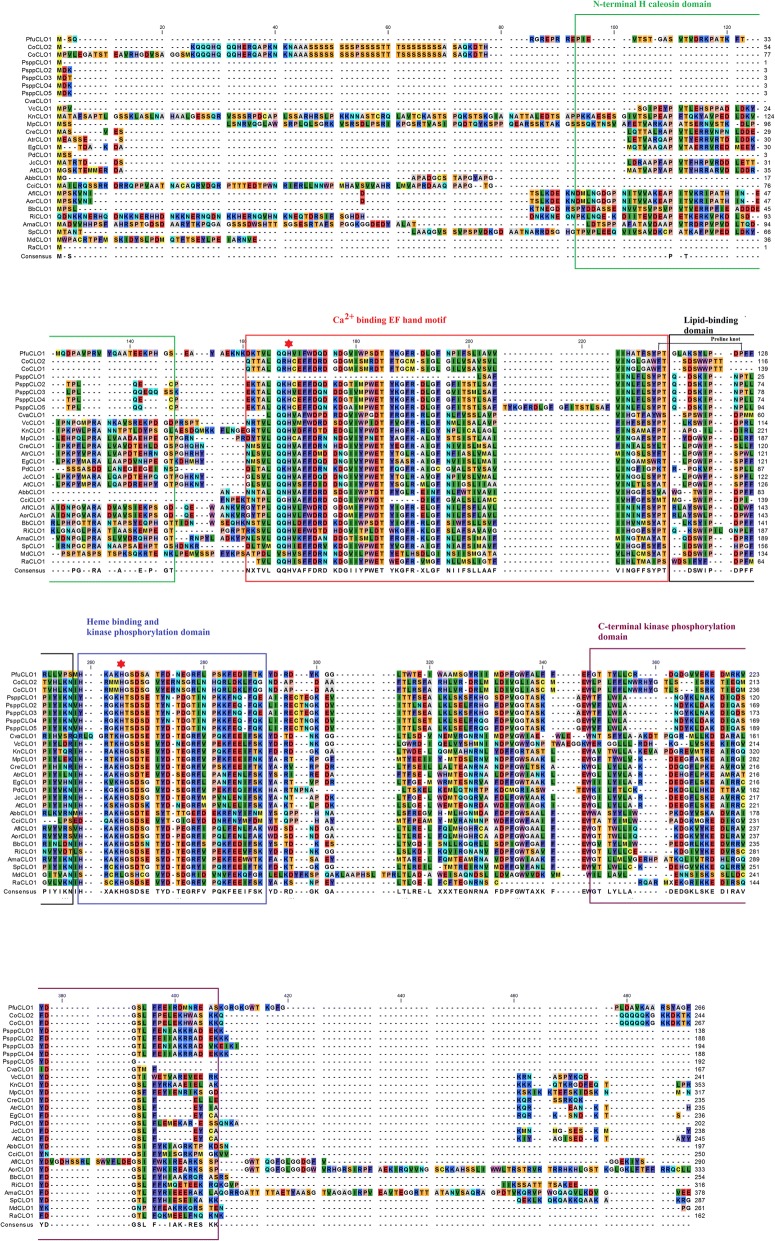
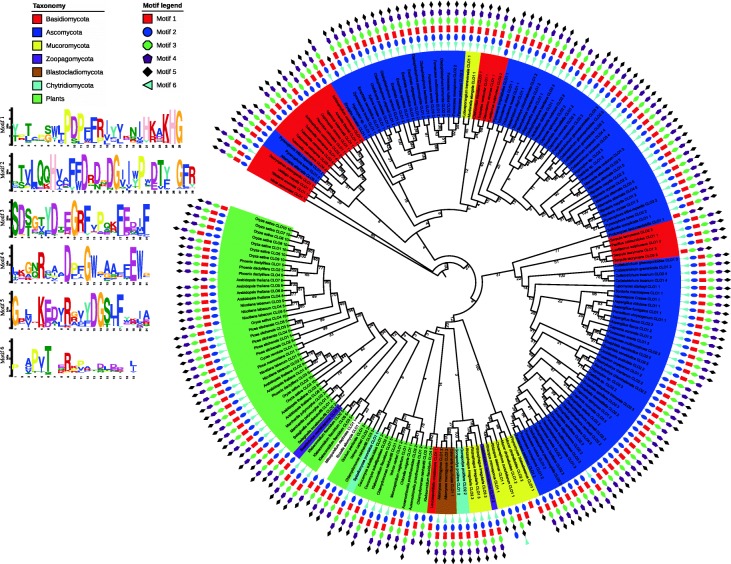
Results: CLO/PXG-like genes are distributed across the full range of fungal groups from the basal clades, Cryptomycota and Microsporidia, to the largest and most complex Dikarya species. However, the genes were only present in 243 out of 844 analysed fungal genomes. CLO/PXG-like genes have been retained in many pathogenic or parasitic fungi that have undergone considerable genomic and structural simplification, indicating that they have important functions in these species. Structural and functional analyses demonstrate that CLO/PXGs are multifunctional proteins closely related to similar proteins found in all major taxa of the Chlorophyte Division of the Viridiplantae. Transcriptome and physiological data show that fungal CLO/PXG-like genes have complex patterns of developmental and tissue-specific expression and are upregulated in response to a range of biotic and abiotic stresses as well as participating in key metabolic and developmental processes such as lipid metabolism, signalling, reproduction and pathogenesis. Biochemical data also reveal that the Aspergillus flavus CLO/PXG has specific functions in sporulation and aflatoxin production as well as playing roles in lipid droplet function.
Conclusions: In contrast to plants, CLO/PXGs only occur in about 30% of sequenced fungal genomes but are present in all major taxa. Fungal CLO/PXGs have similar but not identical roles to those in plants, including stress-related oxylipin signalling, lipid metabolism, reproduction and pathogenesis. While the presence of CLO/PXG orthologs in all plant genomes sequenced to date would suggest that they have core housekeeping functions in plants, the selective loss of CLO/PXGs in many fungal genomes suggests more restricted functions in fungi as accessory genes useful in particular environments or niches. We suggest an ancient origin of CLO/PXG-like genes in the 'last eukaryotic common ancestor' (LECA) and their subsequent loss in ancestors of the Metazoa, after the latter had diverged from the ancestral fungal lineage.
Keywords: Caleosin; Evolution; Fungi; Lipid droplets; Oxylipins; Peroxygenase; Stress responses; Viridiplantae.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Consent for publication
All required consents have been received.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures


Similar articles
-
Evolutionary and genomic analysis of the caleosin/peroxygenase (CLO/PXG) gene/protein families in the Viridiplantae.PLoS One. 2018 May 17;13(5):e0196669. doi: 10.1371/journal.pone.0196669. eCollection 2018. PLoS One. 2018. PMID: 29771926 Free PMC article.
-
Caleosin/peroxygenases: multifunctional proteins in plants.Ann Bot. 2023 Apr 4;131(3):387-409. doi: 10.1093/aob/mcad001. Ann Bot. 2023. PMID: 36656070 Free PMC article. Review.
-
A Caleosin-Like Protein with Peroxygenase Activity Mediates Aspergillus flavus Development, Aflatoxin Accumulation, and Seed Infection.Appl Environ Microbiol. 2015 Sep;81(18):6129-44. doi: 10.1128/AEM.00867-15. Epub 2015 Jun 26. Appl Environ Microbiol. 2015. PMID: 26116672 Free PMC article.
-
Roles of a membrane-bound caleosin and putative peroxygenase in biotic and abiotic stress responses in Arabidopsis.Plant Physiol Biochem. 2009 Sep;47(9):796-806. doi: 10.1016/j.plaphy.2009.04.005. Epub 2009 May 9. Plant Physiol Biochem. 2009. PMID: 19467604
-
The Fungal Kingdom: diverse and essential roles in earth's ecosystem: This report is based on a colloquium, sponsored by the American Academy of Microbiology, convened November 2–4, 2007 in Tucson, Arizona.Washington (DC): American Society for Microbiology; 2008. Washington (DC): American Society for Microbiology; 2008. PMID: 32687285 Free Books & Documents. Review.
Cited by
-
Lessons on fruiting body morphogenesis from genomes and transcriptomes of Agaricomycetes.Stud Mycol. 2023 Jul;104:1-85. doi: 10.3114/sim.2022.104.01. Epub 2023 Jan 31. Stud Mycol. 2023. PMID: 37351542 Free PMC article.
-
3D-reconstructions of zygospores in Zygnema vaginatum (Charophyta) reveal details of cell wall formation, suggesting adaptations to extreme habitats.Physiol Plant. 2023 Jul-Aug;175(4):e13988. doi: 10.1111/ppl.13988. Physiol Plant. 2023. PMID: 37616005 Free PMC article.
-
Genomic Analysis of the Caleosin Family in Theaceae Reveals Lineagespecific Evolutionary Patterns.Curr Protein Pept Sci. 2025;26(2):139-155. doi: 10.2174/0113892037321073240828051039. Curr Protein Pept Sci. 2025. PMID: 39323337
-
Transcriptome Analysis of Bread Wheat Genotype KRL3-4 Provides a New Insight Into Regulatory Mechanisms Associated With Sodicity (High pH) Tolerance.Front Genet. 2022 Feb 9;12:782366. doi: 10.3389/fgene.2021.782366. eCollection 2021. Front Genet. 2022. PMID: 35222517 Free PMC article.
-
Degradation of Lipid Droplets in Plants and Algae-Right Time, Many Paths, One Goal.Front Plant Sci. 2020 Sep 9;11:579019. doi: 10.3389/fpls.2020.579019. eCollection 2020. Front Plant Sci. 2020. PMID: 33014002 Free PMC article. Review.
References
-
- Hanano A, Alkara M, Almousally I, Shaban M, Rahman F, Hassan M, Murphy DJ. The Peroxygenase activity of the Aspergillus flavus Caleosin, AfPXG, modulates the biosynthesis of aflatoxins and their trafficking and extracellular secretion via lipid droplets. Front Microbiol. 2018;9:158. doi: 10.3389/fmicb.2018.00158. - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
