

The CMG Helicase Bypasses DNA-Protein Cross-Links to Facilitate Their Repair
- PMID: 30595447
- PMCID: PMC6475077
- DOI: 10.1016/j.cell.2018.10.053
The CMG Helicase Bypasses DNA-Protein Cross-Links to Facilitate Their Repair
Abstract
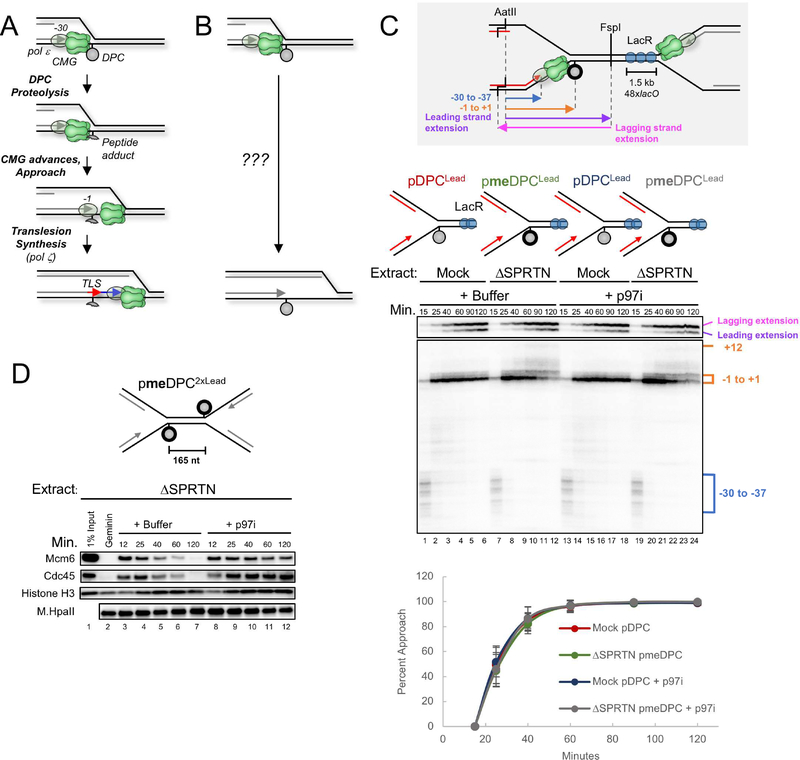
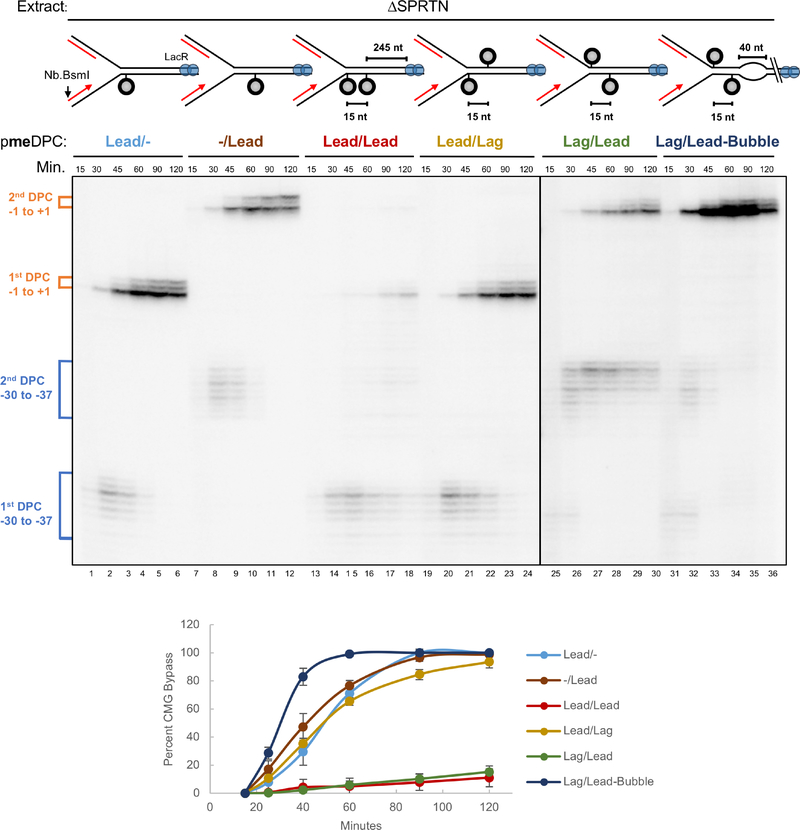
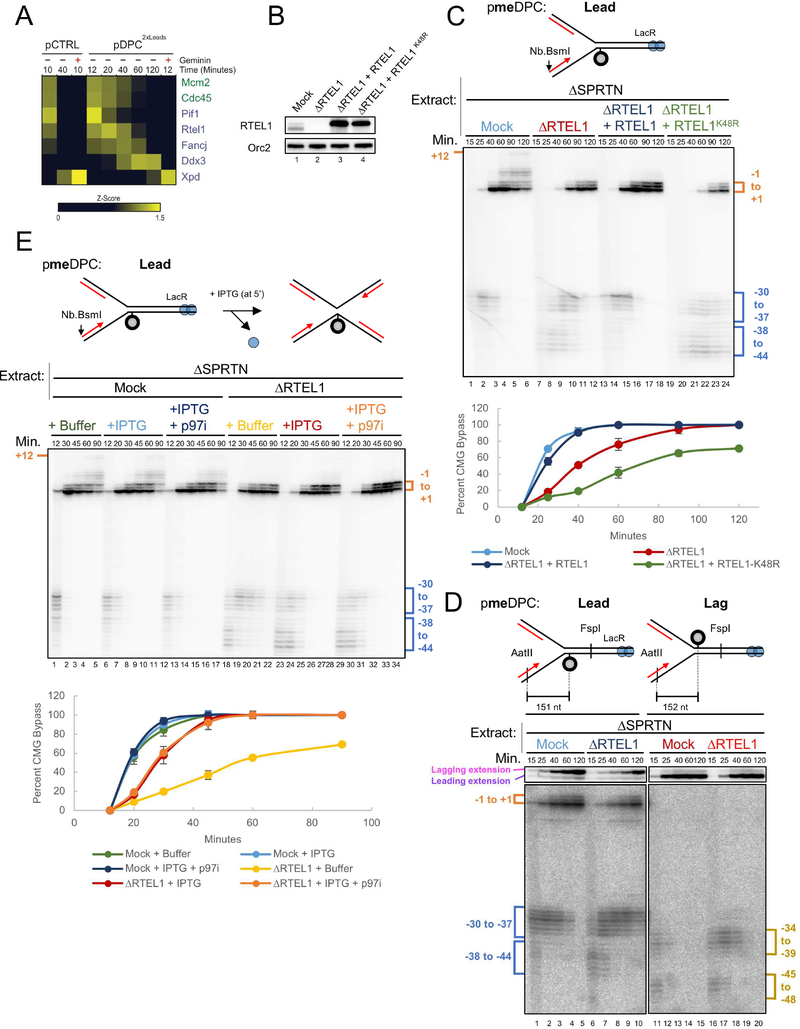
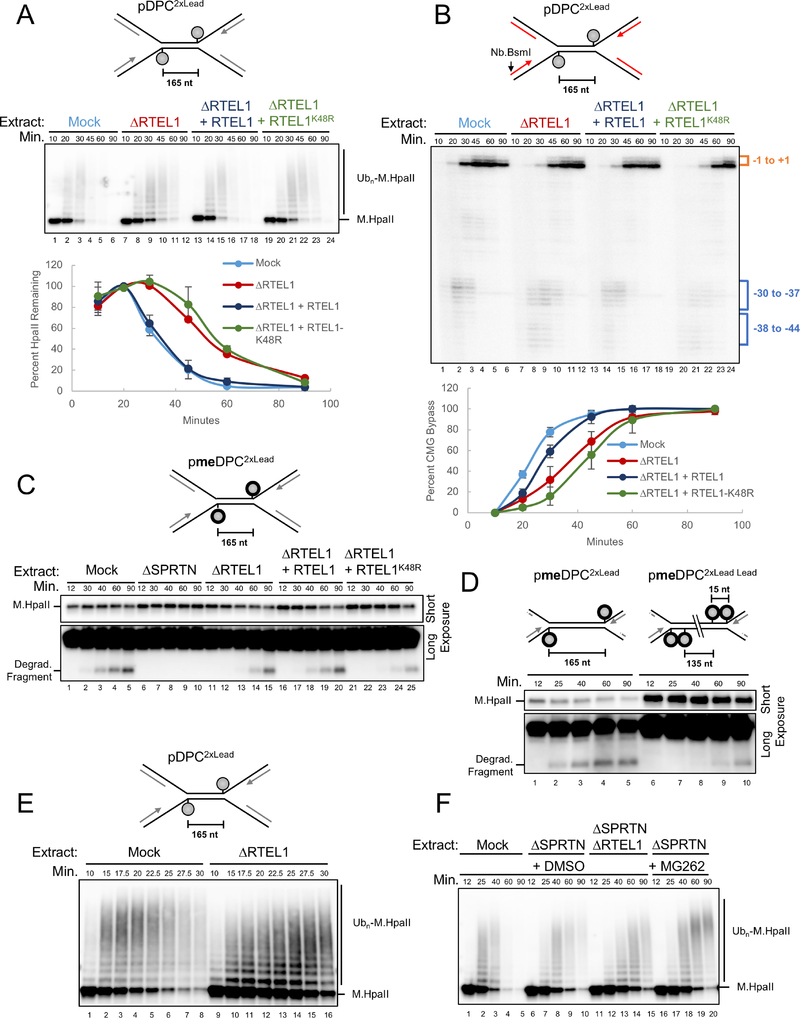
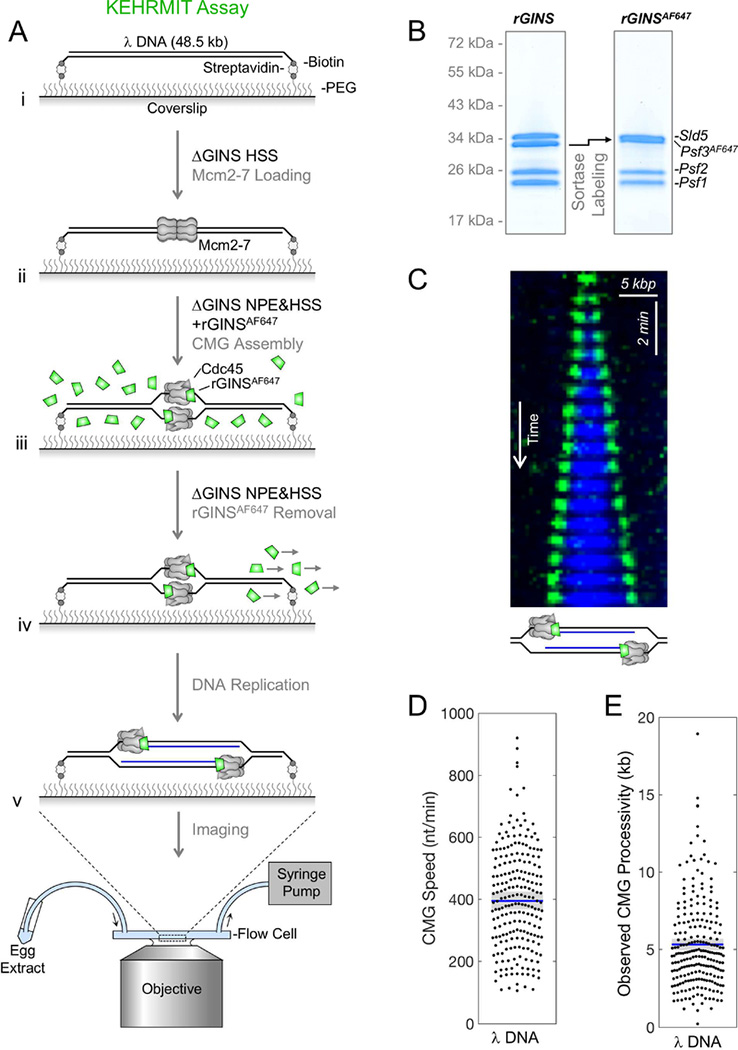
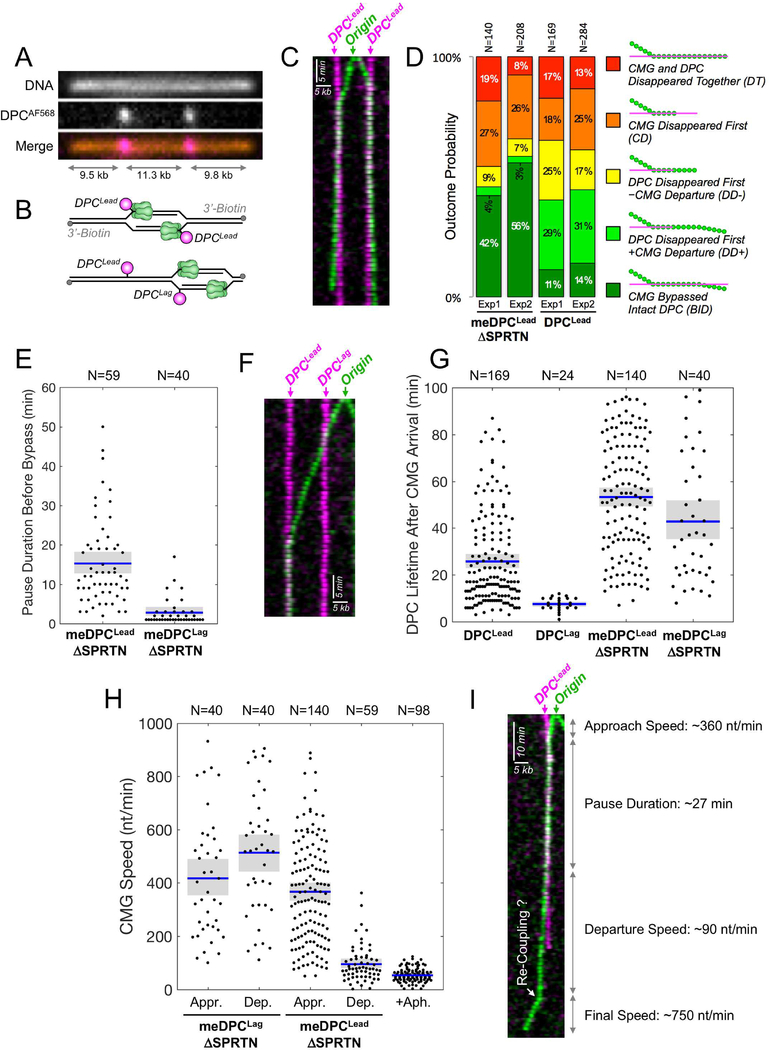
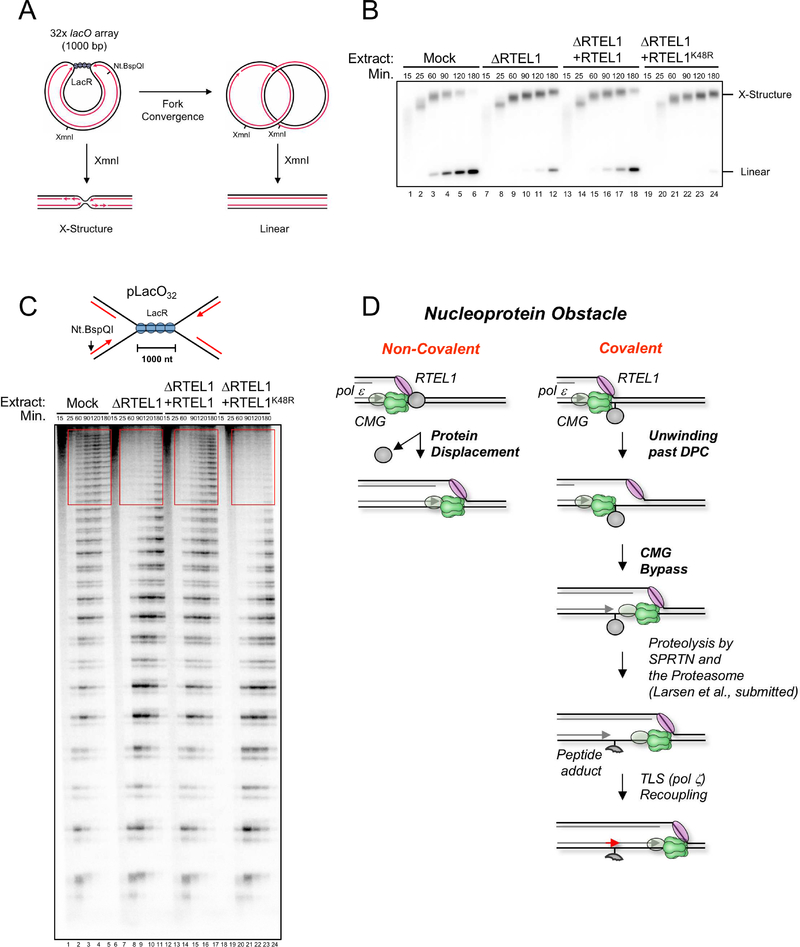
Covalent DNA-protein cross-links (DPCs) impede replication fork progression and threaten genome integrity. Using Xenopus egg extracts, we previously showed that replication fork collision with DPCs causes their proteolysis, followed by translesion DNA synthesis. We show here that when DPC proteolysis is blocked, the replicative DNA helicase CMG (CDC45, MCM2-7, GINS), which travels on the leading strand template, bypasses an intact leading strand DPC. Single-molecule imaging reveals that GINS does not dissociate from CMG during bypass and that CMG slows dramatically after bypass, likely due to uncoupling from the stalled leading strand. The DNA helicase RTEL1 facilitates bypass, apparently by generating single-stranded DNA beyond the DPC. The absence of RTEL1 impairs DPC proteolysis, suggesting that CMG must bypass the DPC to enable proteolysis. Our results suggest a mechanism that prevents inadvertent CMG destruction by DPC proteases, and they reveal CMG's remarkable capacity to overcome obstacles on its translocation strand.
Keywords: CMG; DNA-protein cross-links; KEHRMIT; Xenopus laevis egg extracts; bulky lesion bypass; eukaryotic DNA replication; eukaryotic replicative helicase; helicase uncoupling; replication-coupled DNA repair; single-molecule approach.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures
References
-
- Bochman ML, and Schwacha A (2008). The Mcm2–7 complex has in vitro helicase activity. Mol Cell 31, 287–293. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
