

Efferocytosis Fuels Requirements of Fatty Acid Oxidation and the Electron Transport Chain to Polarize Macrophages for Tissue Repair
- PMID: 30595481
- PMCID: PMC6471613
- DOI: 10.1016/j.cmet.2018.12.004
Efferocytosis Fuels Requirements of Fatty Acid Oxidation and the Electron Transport Chain to Polarize Macrophages for Tissue Repair
Abstract
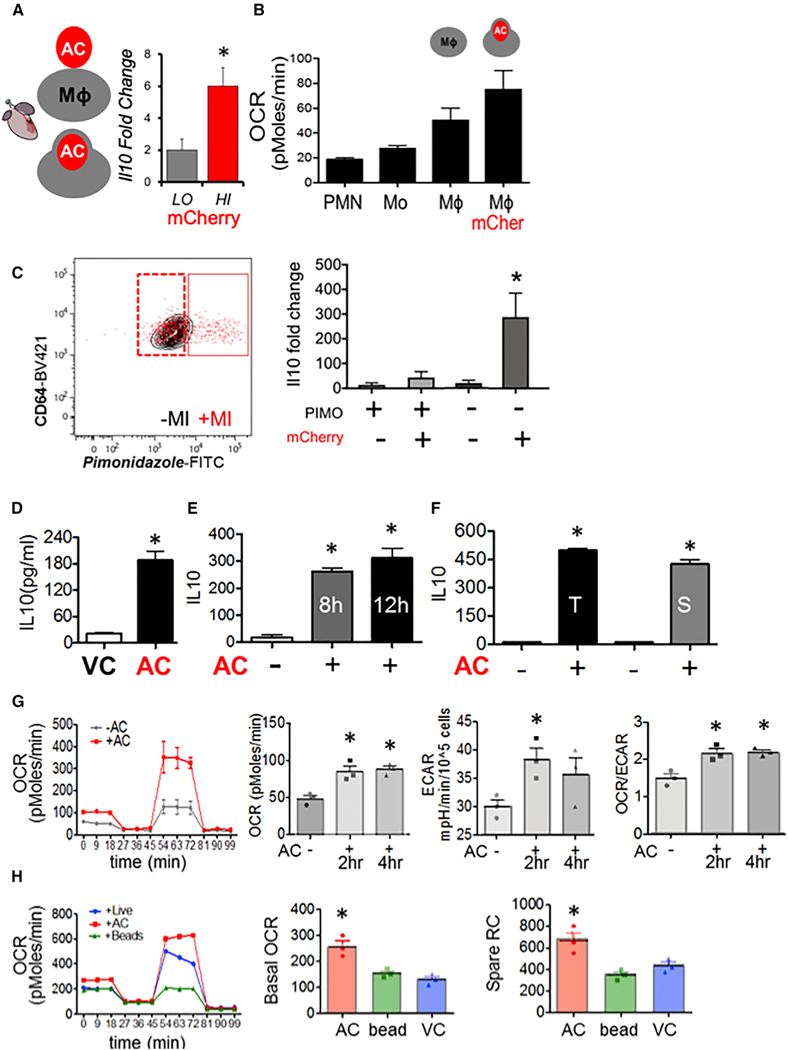
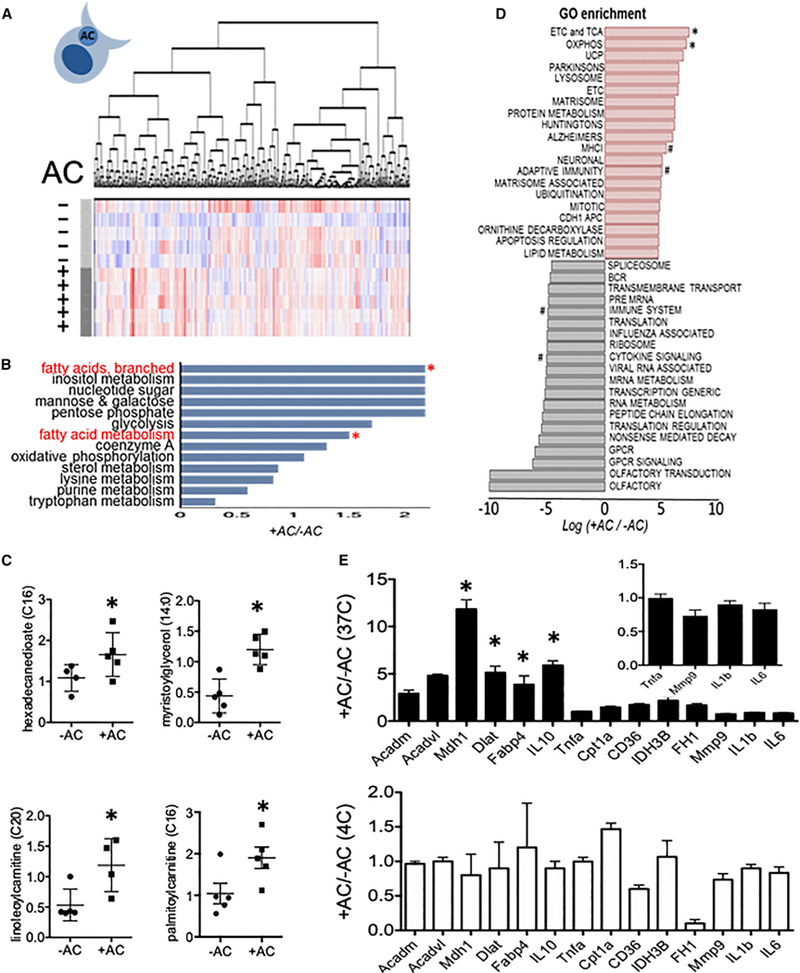
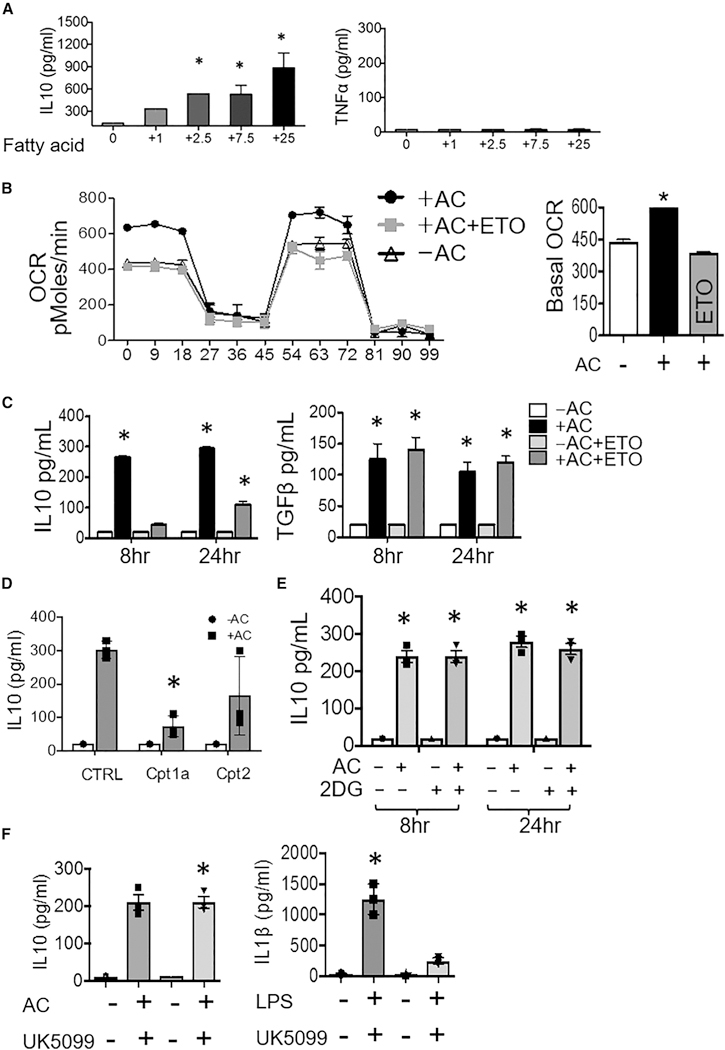
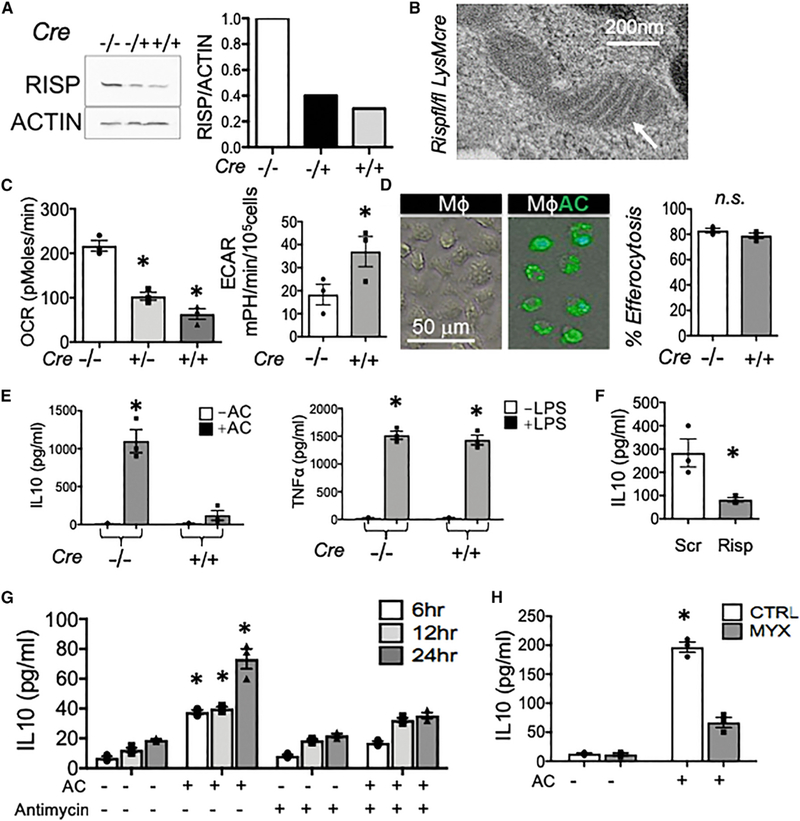
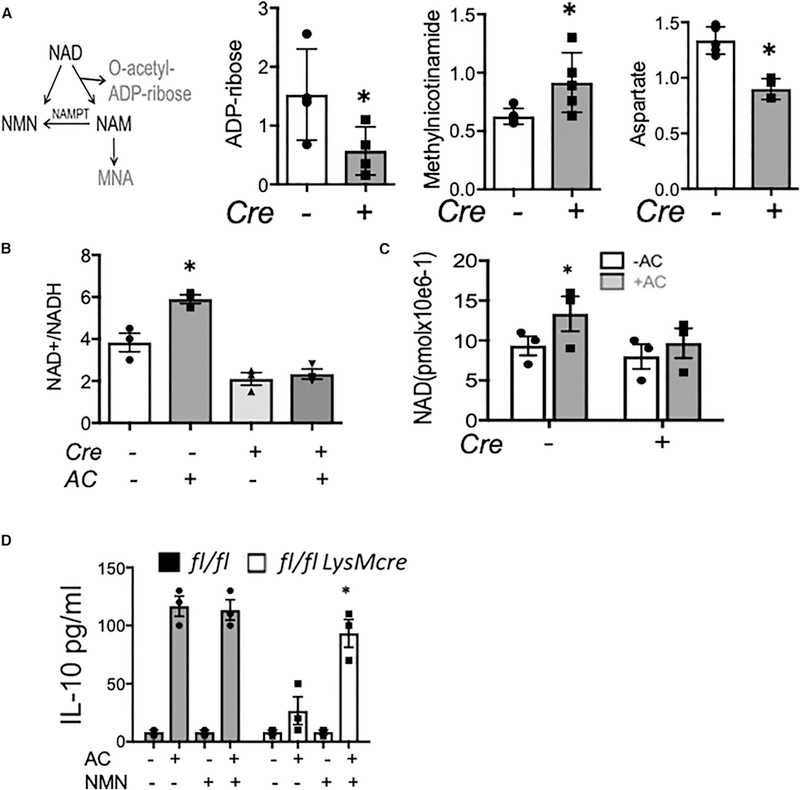
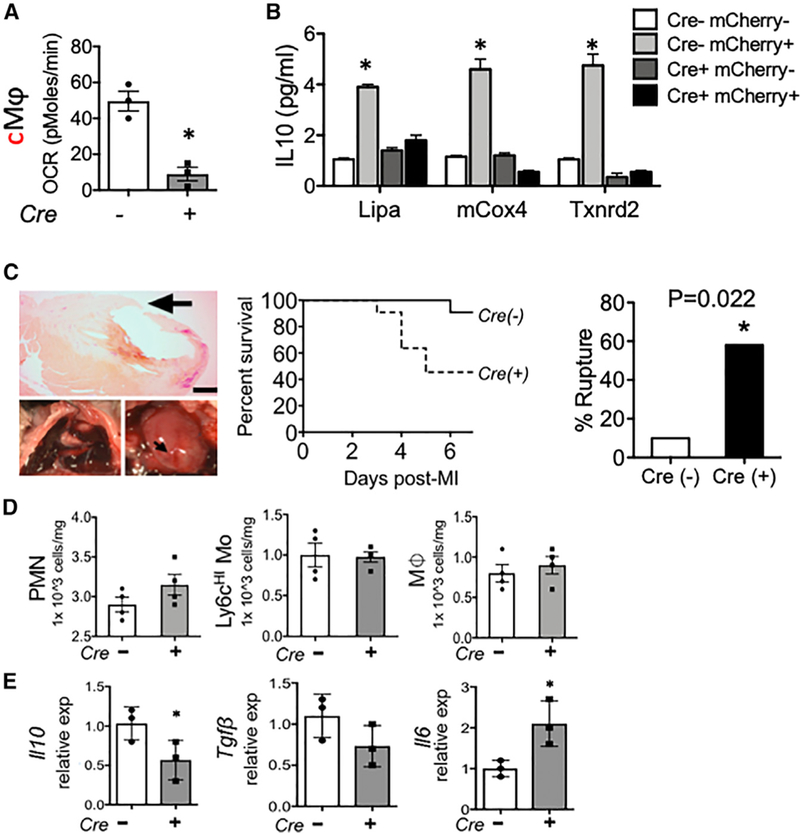
During wound injury, efferocytosis fills the macrophage with a metabolite load nearly equal to the phagocyte itself. A timely question pertains to how metabolic phagocytic signaling regulates the signature anti-inflammatory macrophage response. Here we report the metabolome of activated macrophages during efferocytosis to reveal an interleukin-10 (IL-10) cytokine escalation that was independent of glycolysis yet bolstered by apoptotic cell fatty acids and mitochondrial β-oxidation, the electron transport chain, and heightened coenzyme NAD+. Loss of IL-10 due to mitochondrial complex III defects was remarkably rescued by adding NAD+ precursors. This activated a SIRTUIN1 signaling cascade, largely independent of ATP, that culminated in activation of IL-10 transcription factor PBX1. Il-10 activation by the respiratory chain was also important in vivo, as efferocyte mitochondrial dysfunction led to cardiac rupture after myocardial injury. These findings highlight a new paradigm whereby macrophages leverage efferocytic metabolites and electron transport for anti-inflammatory reprogramming that culminates in organ repair.
Keywords: efferocytosis; immunometabolism; macrophage; wound healing.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures

Comment in
-
Death Eaters Rely on Metabolic Signaling to Wield Anti-inflammatory Responses.Cell Metab. 2019 Feb 5;29(2):234-236. doi: 10.1016/j.cmet.2019.01.010. Cell Metab. 2019. PMID: 30726754
References
-
- Amat R, Planavila A, Chen SL, Iglesias R, Giralt M, and Villarroya F (2009). SIRT1 controls the transcription of the peroxisome proliferator-activated receptor-gamma Co-activator-1alpha (PGC-1alpha) gene in skeletal muscle through the PGC-1alpha autoregulatory loop and interaction with MyoD. J. Biol. Chem. 284, 21872–21880. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
