

A Phytophthora Effector Suppresses Trans-Kingdom RNAi to Promote Disease Susceptibility
- PMID: 30595554
- PMCID: PMC9208300
- DOI: 10.1016/j.chom.2018.11.007
A Phytophthora Effector Suppresses Trans-Kingdom RNAi to Promote Disease Susceptibility
Abstract
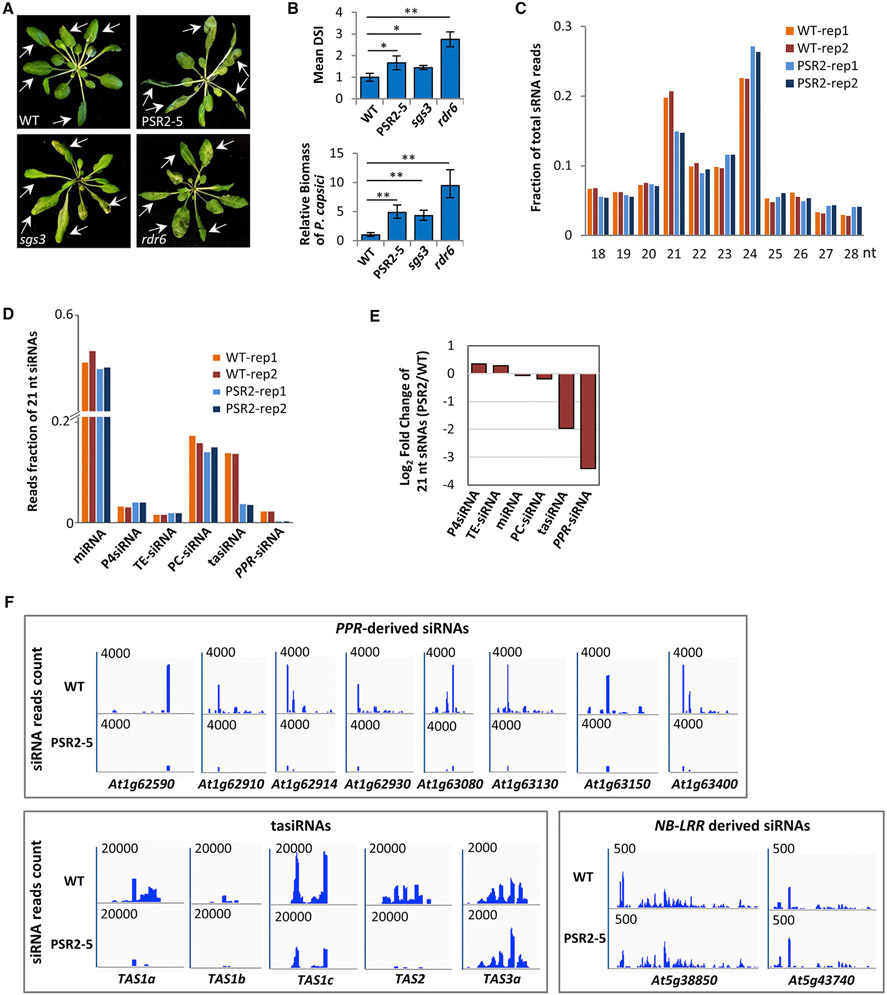
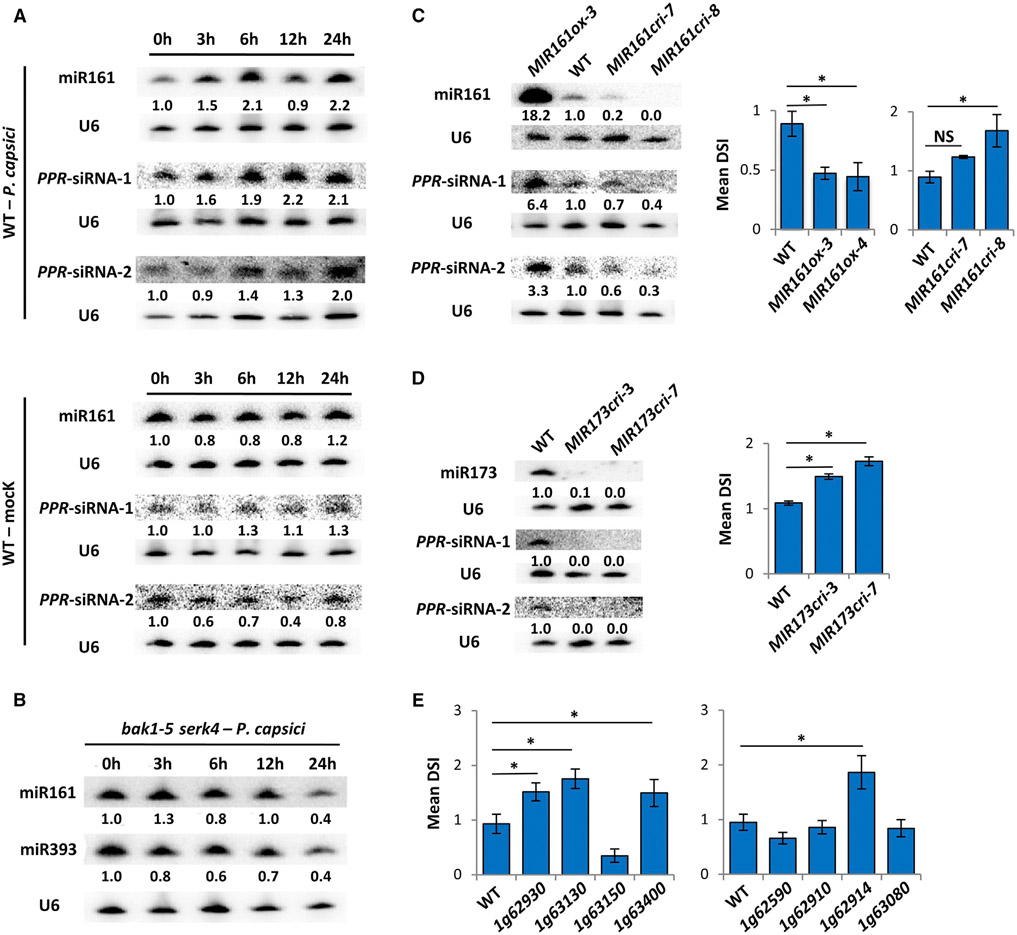
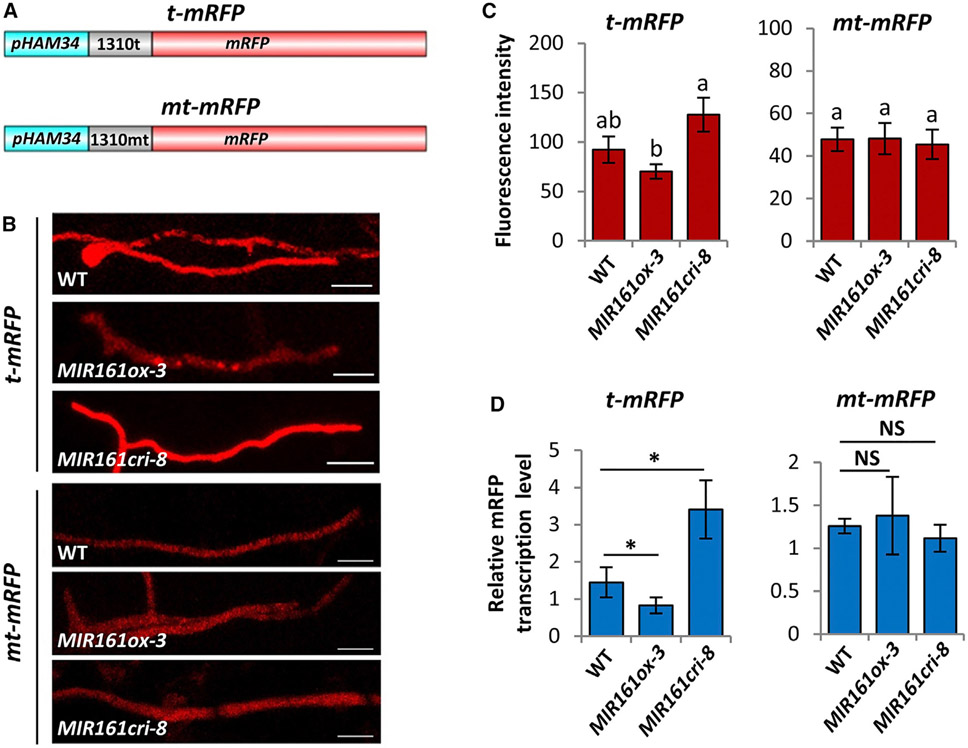
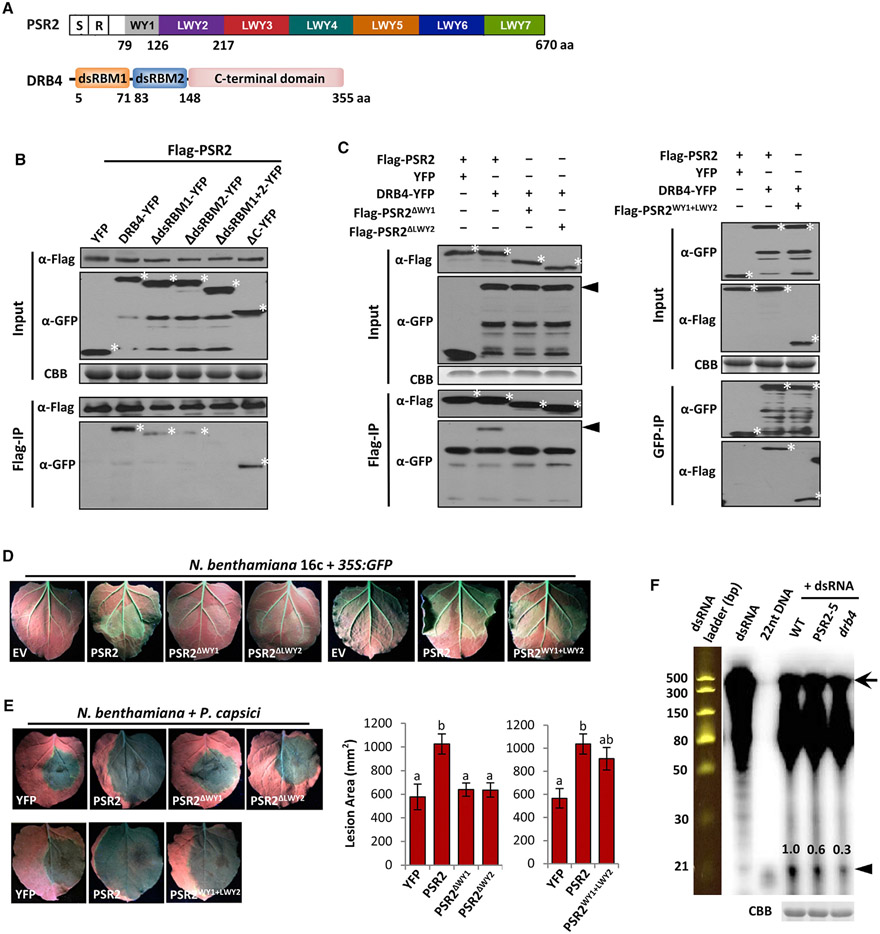
RNA silencing (RNAi) has a well-established role in anti-viral immunity in plants. The destructive eukaryotic pathogen Phytophthora encodes suppressors of RNAi (PSRs), which enhance plant susceptibility. However, the role of small RNAs in defense against eukaryotic pathogens is unclear. Here, we show that Phytophthora infection of Arabidopsis leads to increased production of a diverse pool of secondary small interfering RNAs (siRNAs). Instead of regulating endogenous plant genes, these siRNAs are found in extracellular vesicles and likely silence target genes in Phytophthora during natural infection. Introduction of a plant siRNA in Phytophthora leads to developmental deficiency and abolishes virulence, while Arabidopsis mutants defective in secondary siRNA biogenesis are hypersusceptible. Notably, Phytophthora effector PSR2 specifically inhibits secondary siRNA biogenesis in Arabidopsis and promotes infection. These findings uncover the role of siRNAs as antimicrobial agents against eukaryotic pathogens and highlight a defense/counter-defense arms race centered on trans-kingdom gene silencing between hosts and pathogens.
Keywords: RNA interference; RNA-silencing suppressor; host-induced gene silencing; microRNA; pathogenesis; plant immunity; secondary small RNAs; virulence factor.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures


Comment in
-
Second to None: Plant Secondary siRNAs as Defensive Agents against Phytophthora.Cell Host Microbe. 2019 Jan 9;25(1):7-9. doi: 10.1016/j.chom.2018.12.007. Cell Host Microbe. 2019. PMID: 30629921
Similar articles
-
Phytophthora suppressor of RNA silencing 2 is a conserved RxLR effector that promotes infection in soybean and Arabidopsis thaliana.Mol Plant Microbe Interact. 2014 Dec;27(12):1379-89. doi: 10.1094/MPMI-06-14-0190-R. Mol Plant Microbe Interact. 2014. PMID: 25387135
-
Small RNAs - Big Players in Plant-Microbe Interactions.Cell Host Microbe. 2019 Aug 14;26(2):173-182. doi: 10.1016/j.chom.2019.07.021. Cell Host Microbe. 2019. PMID: 31415750 Review.
-
Virus infection triggers widespread silencing of host genes by a distinct class of endogenous siRNAs in Arabidopsis.Proc Natl Acad Sci U S A. 2014 Oct 7;111(40):14613-8. doi: 10.1073/pnas.1407131111. Epub 2014 Sep 8. Proc Natl Acad Sci U S A. 2014. PMID: 25201959 Free PMC article.
-
Phytophthora effector targets a novel component of small RNA pathway in plants to promote infection.Proc Natl Acad Sci U S A. 2015 May 5;112(18):5850-5. doi: 10.1073/pnas.1421475112. Epub 2015 Apr 20. Proc Natl Acad Sci U S A. 2015. PMID: 25902521 Free PMC article.
-
Filamentous pathogen effectors interfering with small RNA silencing in plant hosts.Curr Opin Microbiol. 2016 Aug;32:1-6. doi: 10.1016/j.mib.2016.04.003. Epub 2016 Apr 20. Curr Opin Microbiol. 2016. PMID: 27104934 Review.
Cited by
-
Challenges and Opportunities Arising from Host-Botrytis cinerea Interactions to Outline Novel and Sustainable Control Strategies: The Key Role of RNA Interference.Int J Mol Sci. 2024 Jun 20;25(12):6798. doi: 10.3390/ijms25126798. Int J Mol Sci. 2024. PMID: 38928507 Free PMC article. Review.
-
Exchange of Small Regulatory RNAs between Plants and Their Pests.Plant Physiol. 2020 Jan;182(1):51-62. doi: 10.1104/pp.19.00931. Epub 2019 Oct 21. Plant Physiol. 2020. PMID: 31636103 Free PMC article.
-
Action Mechanisms of Effectors in Plant-Pathogen Interaction.Int J Mol Sci. 2022 Jun 17;23(12):6758. doi: 10.3390/ijms23126758. Int J Mol Sci. 2022. PMID: 35743201 Free PMC article. Review.
-
New Insights on the Integrated Management of Plant Diseases by RNA Strategies: Mycoviruses and RNA Interference.Int J Mol Sci. 2022 Aug 17;23(16):9236. doi: 10.3390/ijms23169236. Int J Mol Sci. 2022. PMID: 36012499 Free PMC article. Review.
-
Identification of genetic loci in lettuce mediating quantitative resistance to fungal pathogens.Theor Appl Genet. 2022 Jul;135(7):2481-2500. doi: 10.1007/s00122-022-04129-5. Epub 2022 Jun 8. Theor Appl Genet. 2022. PMID: 35674778 Free PMC article.
References
-
- Adenot X, Elmayan T, Lauressergues D, Boutet S, Bouche N, Gasciolli V, and Vaucheret H (2006). DRB4-dependent TAS3 trans-acting siRNAs control leaf morphology through AGO7. Curr. Biol 16, 927–932. - PubMed
-
- An Q, Ehlers K, Kogel KH, van Bel AJ, and Huckelhoven R (2006a). Multivesicular compartments proliferate in susceptible and resistant MLA12-barley leaves in response to infection by the biotrophic powdery mildew fungus. New Phytol. 172, 563–576. - PubMed
-
- An Q, Huckelhoven R, Kogel KH, and van Bel AJ (2006b). Multivesicular bodies participate in a cell wall-associated defence response in barley leaves attacked by the pathogenic powdery mildew fungus. Cell Microbiol. 8, 1009–1019. - PubMed
-
- Axtell MJ (2013). Classification and comparison of small RNAs from plants. Annu. Rev. Plant Biol 64, 137–159. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
