

A Comparative Perspective on Brain Regeneration in Amphibians and Teleost Fish
- PMID: 30600647
- PMCID: PMC6618004
- DOI: 10.1002/dneu.22665
A Comparative Perspective on Brain Regeneration in Amphibians and Teleost Fish
Abstract
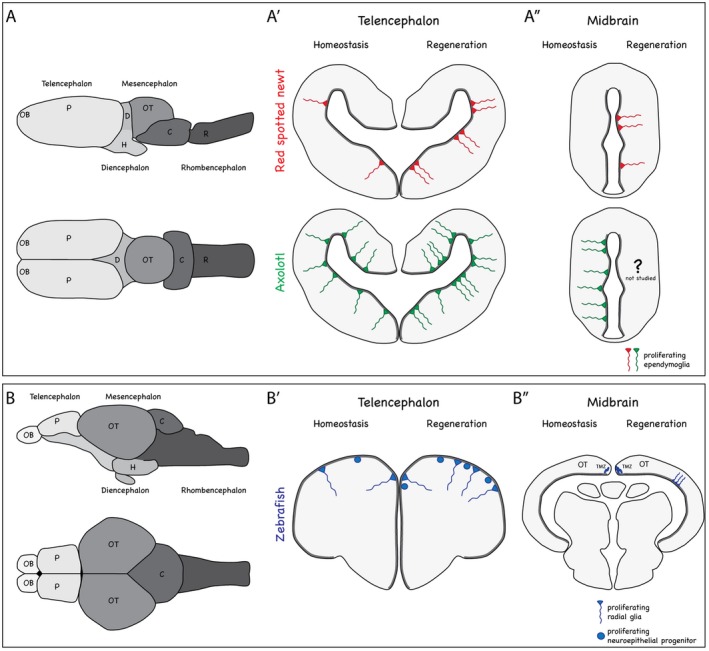
Regeneration of lost cells in the central nervous system, especially the brain, is present to varying degrees in different species. In mammals, neuronal cell death often leads to glial cell hypertrophy, restricted proliferation, and formation of a gliotic scar, which prevents neuronal regeneration. Conversely, amphibians such as frogs and salamanders and teleost fish possess the astonishing capacity to regenerate lost cells in several regions of their brains. While frogs lose their regenerative abilities after metamorphosis, teleost fish and salamanders are known to possess regenerative competence even throughout adulthood. In the last decades, substantial progress has been made in our understanding of the cellular and molecular mechanisms of brain regeneration in amphibians and fish. But how similar are the means of brain regeneration in these different species? In this review, we provide an overview of common and distinct aspects of brain regeneration in frog, salamander, and teleost fish species: from the origin of regenerated cells to the functional recovery of behaviors.
Keywords: amphibian; brain; regeneration; teleost fish.
© 2019 The Authors. Developmental Neurobiology Published by Wiley Periodicals, Inc.
Conflict of interest statement
The authors indicate no potential conflicts of interests.
Figures
Similar articles
-
Organ regeneration evolved in fish and amphibians in relation to metamorphosis: Speculations on a post-embryonic developmental process lost in amniotes after the water to land transition.Ann Anat. 2019 Mar;222:114-119. doi: 10.1016/j.aanat.2018.12.005. Epub 2018 Dec 20. Ann Anat. 2019. PMID: 30580055 Review.
-
The African clawed frog Xenopus laevis: A model organism to study regeneration of the central nervous system.Neurosci Lett. 2017 Jun 23;652:82-93. doi: 10.1016/j.neulet.2016.09.054. Epub 2016 Sep 29. Neurosci Lett. 2017. PMID: 27693567 Review.
-
Spinal cord repair in regeneration-competent vertebrates: adult teleost fish as a model system.Brain Res Rev. 2011 Jun 24;67(1-2):73-93. doi: 10.1016/j.brainresrev.2010.11.001. Epub 2010 Nov 6. Brain Res Rev. 2011. PMID: 21059372 Review.
-
Astrocytes as gate-keepers in optic nerve regeneration--a mini-review.Comp Biochem Physiol A Mol Integr Physiol. 2009 Feb;152(2):135-8. doi: 10.1016/j.cbpa.2008.09.026. Epub 2008 Oct 2. Comp Biochem Physiol A Mol Integr Physiol. 2009. PMID: 18930160 Review.
-
The promise of perfect adult tissue repair and regeneration in mammals: Learning from regenerative amphibians and fish.Bioessays. 2014 Sep;36(9):861-71. doi: 10.1002/bies.201300144. Epub 2014 Jul 9. Bioessays. 2014. PMID: 25043537 Review.
Cited by
-
Study of Natural Longlife Juvenility and Tissue Regeneration in Caudate Amphibians and Potential Application of Resulting Data in Biomedicine.J Dev Biol. 2021 Jan 18;9(1):2. doi: 10.3390/jdb9010002. J Dev Biol. 2021. PMID: 33477527 Free PMC article. Review.
-
ERK-activated CK-2 triggers blastema formation during appendage regeneration.Sci Adv. 2024 Mar 22;10(12):eadk8331. doi: 10.1126/sciadv.adk8331. Epub 2024 Mar 20. Sci Adv. 2024. PMID: 38507478 Free PMC article.
-
Radial glia and radial glia-like cells: Their role in neurogenesis and regeneration.Front Neurosci. 2022 Nov 16;16:1006037. doi: 10.3389/fnins.2022.1006037. eCollection 2022. Front Neurosci. 2022. PMID: 36466166 Free PMC article. Review.
-
Form and Function of the Vertebrate and Invertebrate Blood-Brain Barriers.Int J Mol Sci. 2021 Nov 9;22(22):12111. doi: 10.3390/ijms222212111. Int J Mol Sci. 2021. PMID: 34829989 Free PMC article. Review.
-
Model systems for regeneration: salamanders.Development. 2019 Jul 22;146(14):dev167700. doi: 10.1242/dev.167700. Development. 2019. PMID: 31332037 Free PMC article. Review.
References
-
- Avila, V.L. and Frye, P.G. (1977) Feeding behavior in the African clawed frog (Xenopus laevis Daudin). Herpetologica, 33, 152–161.
-
- Barbosa, J.S. , Sanchez‐Gonzalez, R. , Di Giaimo, R., Baumgart, E.V. , Theis, F.J. , Götz, M. , et al. (2015) Live imaging of adult neural stem cell behavior in the intact and injured zebrafish brain. Science, 348, 789–793. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
