

Characterization of novel lignocellulose-degrading enzymes from the porcupine microbiome using synthetic metagenomics
- PMID: 30601862
- PMCID: PMC6314593
- DOI: 10.1371/journal.pone.0209221
Characterization of novel lignocellulose-degrading enzymes from the porcupine microbiome using synthetic metagenomics
Abstract
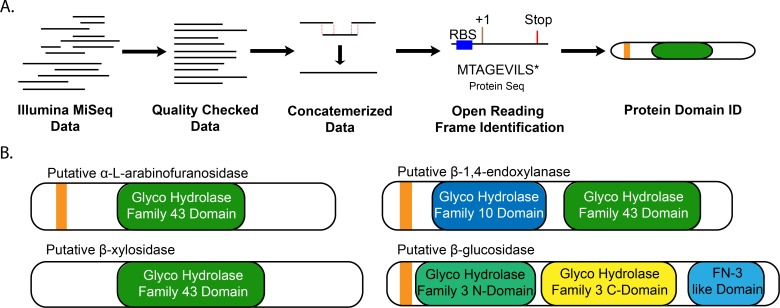
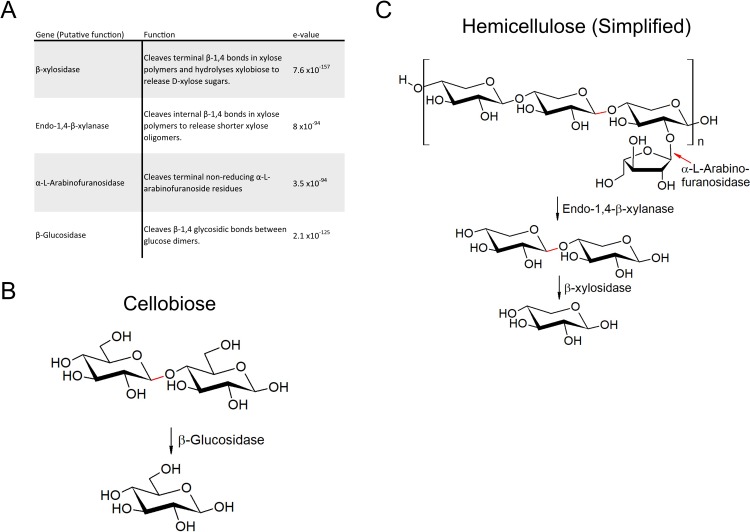
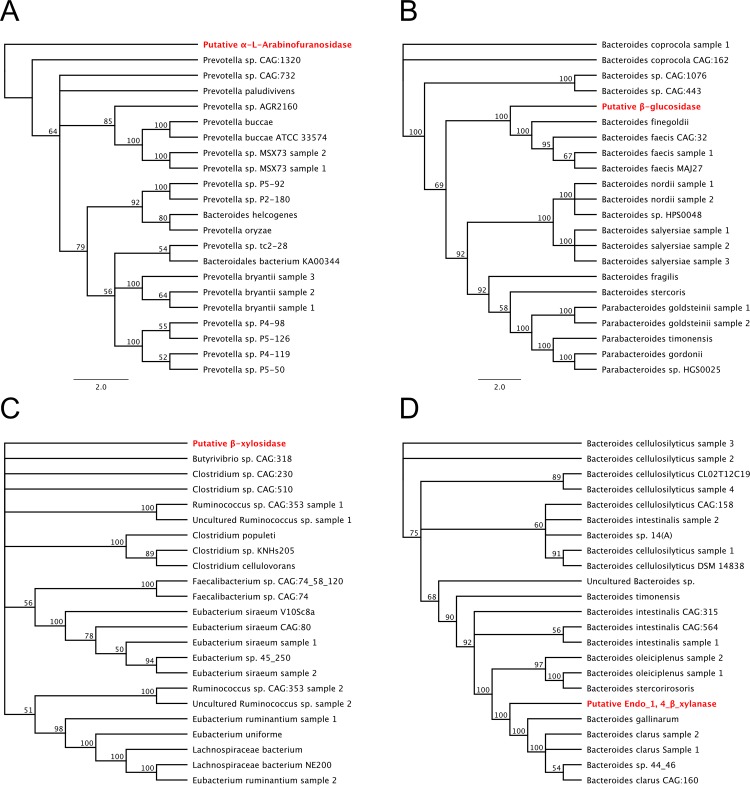
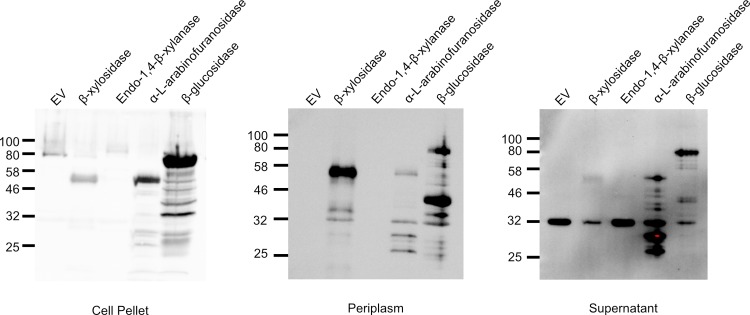
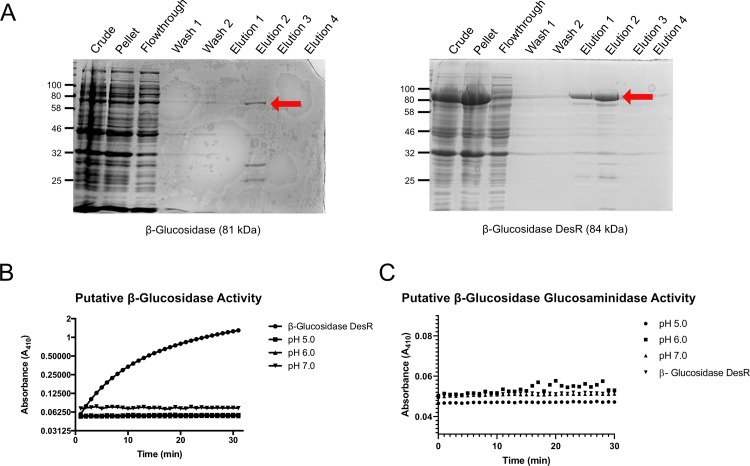
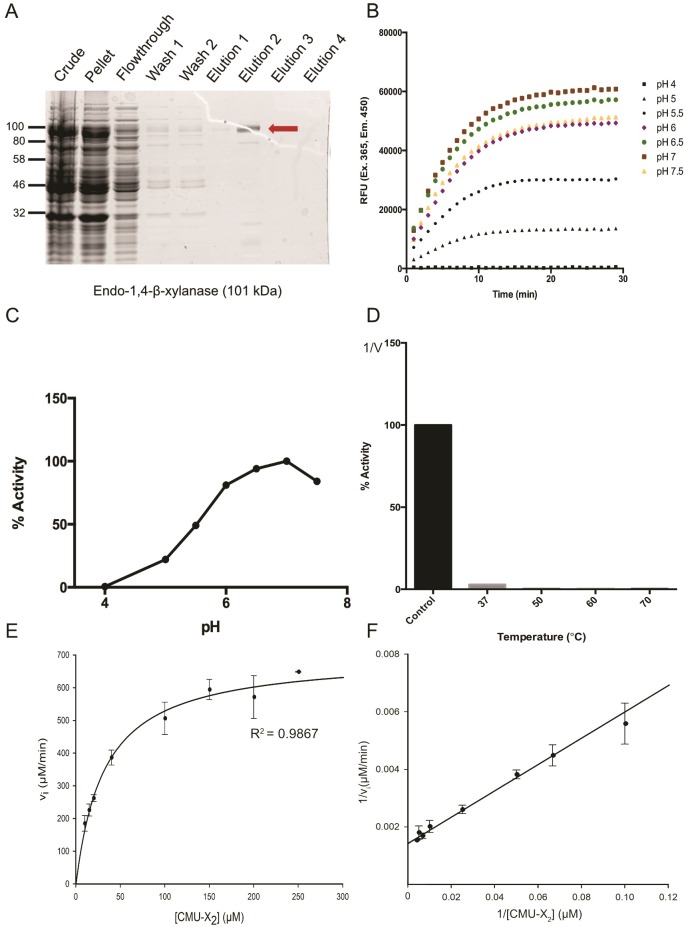
Plant cell walls are composed of cellulose, hemicellulose, and lignin, collectively known as lignocellulose. Microorganisms degrade lignocellulose to liberate sugars to meet metabolic demands. Using a metagenomic sequencing approach, we previously demonstrated that the microbiome of the North American porcupine (Erethizon dorsatum) is replete with genes that could encode lignocellulose-degrading enzymes. Here, we report the identification, synthesis and partial characterization of four novel genes from the porcupine microbiome encoding putative lignocellulose-degrading enzymes: β-glucosidase, α-L-arabinofuranosidase, β-xylosidase, and endo-1,4-β-xylanase. These genes were identified via conserved catalytic domains associated with cellulose- and hemicellulose-degradation. Phylogenetic trees were created for each of these putative enzymes to depict genetic relatedness to known enzymes. Candidate genes were synthesized and cloned into plasmid expression vectors for inducible protein expression and secretion. The putative β-glucosidase fusion protein was efficiently secreted but did not permit Escherichia coli (E. coli) to use cellobiose as a sole carbon source, nor did the affinity purified enzyme cleave p-Nitrophenyl β-D-glucopyranoside (p-NPG) substrate in vitro over a range of physiological pH levels (pH 5-7). The putative hemicellulose-degrading β-xylosidase and α-L-arabinofuranosidase enzymes also lacked in vitro enzyme activity, but the affinity purified endo-1,4-β-xylanase protein cleaved a 6-chloro-4-methylumbelliferyl xylobioside substrate in acidic and neutral conditions, with maximal activity at pH 7. At this optimal pH, KM, Vmax, and kcat were determined to be 32.005 ± 4.72 μM, 1.16x10-5 ± 3.55x10-7 M/s, and 94.72 s-1, respectively. Thus, our pipeline enabled successful identification and characterization of a novel hemicellulose-degrading enzyme from the porcupine microbiome. Progress towards the goal of introducing a complete lignocellulose-degradation pathway into E. coli will be accelerated by combining synthetic metagenomic approaches with functional metagenomic library screening, which can identify novel enzymes unrelated to those found in available databases.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Kumar A, Kushal S, Saraf SA, Singh JS. Microbial biofuels: A solution to carbon emissions and energy crisis. Front Biosci-Landmrk. 2018;23: 1789–1802. - PubMed
-
- Hongzhang C. Biotechnology of Lignocellulose: Theory and Practice. [Internet]. 1st ed Dordrecht: Springer Netherlands; c2014. http://www.springer.com/gp/book/9789400768970. 10.1007/978-94-007-6898-7 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
