

Inhibition of Ongoing Influenza A Virus Replication Reveals Different Mechanisms of RIG-I Activation
- PMID: 30602605
- PMCID: PMC6401434
- DOI: 10.1128/JVI.02066-18
Inhibition of Ongoing Influenza A Virus Replication Reveals Different Mechanisms of RIG-I Activation
Abstract
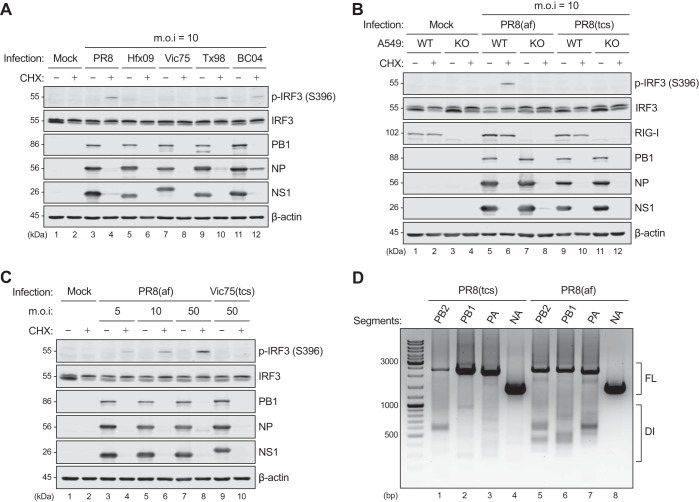
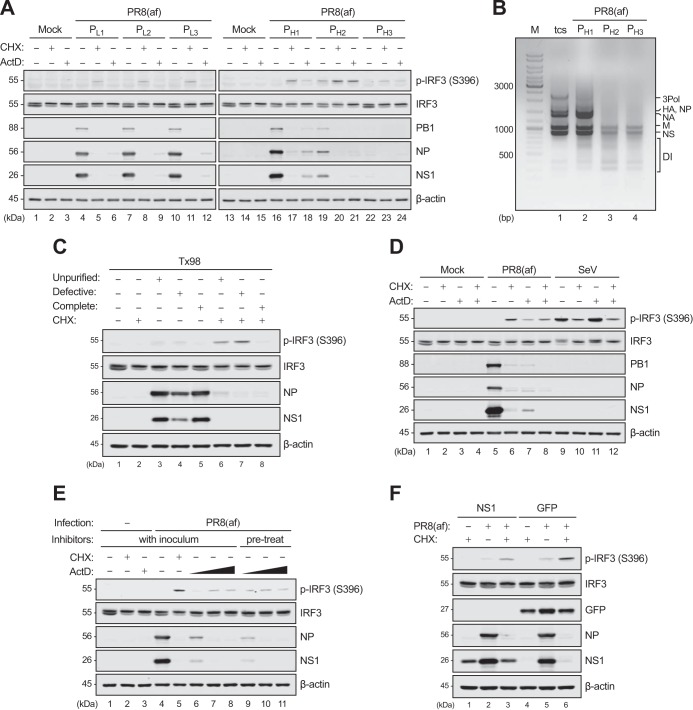
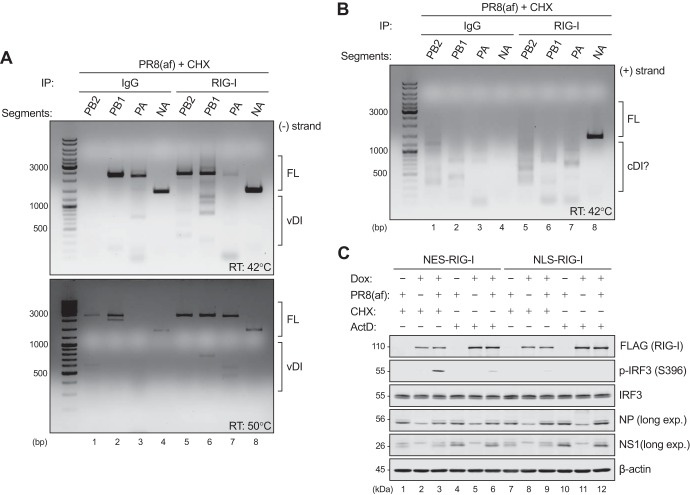
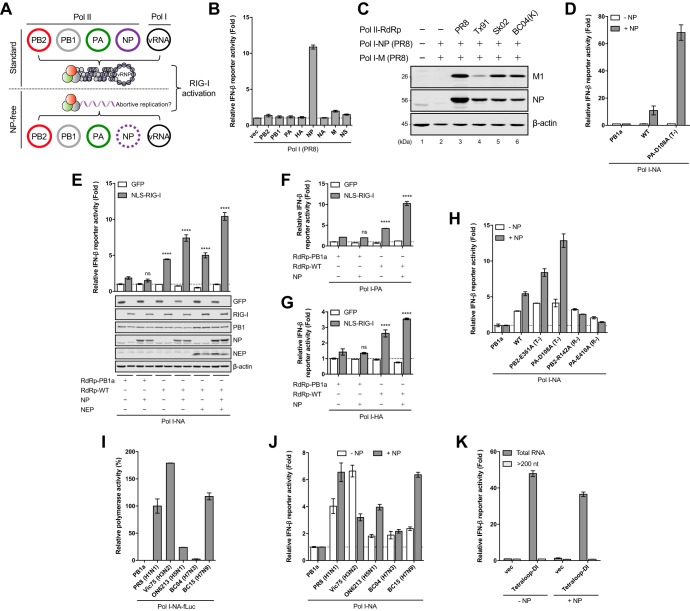
Pattern recognition receptors provide essential nonself immune surveillance within distinct cellular compartments. Retinoic acid-inducible gene I (RIG-I) is one of the primary cytosolic RNA sensors, with an emerging role in the nucleus. It is involved in the spatiotemporal sensing of influenza A virus (IAV) replication, leading to the induction of type I interferons (IFNs). Nonetheless, the physiological viral ligands activating RIG-I during IAV infection remain underexplored. Other than full-length viral genomes, cellular constraints that impede ongoing viral replication likely potentiate an erroneous viral polymerase generating aberrant viral RNA species with RIG-I-activating potential. Here, we investigate the origins of RIG-I-activating viral RNA under two such constraints. Using chemical inhibitors that inhibit continuous viral protein synthesis, we identify the incoming, but not de novo-synthesized, viral defective interfering (DI) genomes contributing to RIG-I activation. In comparison, deprivation of viral nucleoprotein (NP), the key RNA chain elongation factor for the viral polymerase, leads to the production of aberrant viral RNA species activating RIG-I; however, their nature is likely to be distinct from that of DI RNA. Moreover, RIG-I activation in response to NP deprivation is not adversely affected by expression of the nuclear export protein (NEP), which diminishes the generation of a major subset of aberrant viral RNA but facilitates the accumulation of small viral RNA (svRNA). Overall, our results indicate the existence of fundamentally different mechanisms of RIG-I activation under cellular constraints that impede ongoing IAV replication.IMPORTANCE The induction of an IFN response by IAV is mainly mediated by the RNA sensor RIG-I. The physiological RIG-I ligands produced during IAV infection are not fully elucidated. Cellular constraints leading to the inhibition of ongoing viral replication likely potentiate an erroneous viral polymerase producing aberrant viral RNA species activating RIG-I. Here, we demonstrate that RIG-I activation during chemical inhibition of continuous viral protein synthesis is attributable to the incoming DI genomes. Erroneous viral replication driven by NP deprivation promotes the generation of RIG-I-activating aberrant viral RNA, but their nature is likely to be distinct from that of DI RNA. Our results thus reveal distinct mechanisms of RIG-I activation by IAV under cellular constraints impeding ongoing viral replication. A better understanding of RIG-I sensing of IAV infection provides insight into the development of novel interventions to combat influenza virus infection.
Keywords: Influenza A virus; RIG-I; aberrant RNA; defective interfering RNA; innate immunity; interferon.
Copyright © 2019 American Society for Microbiology.
Figures
Similar articles
-
Cytoplasm and Beyond: Dynamic Innate Immune Sensing of Influenza A Virus by RIG-I.J Virol. 2019 Apr 3;93(8):e02299-18. doi: 10.1128/JVI.02299-18. Print 2019 Apr 15. J Virol. 2019. PMID: 30760567 Free PMC article. Review.
-
A Single Amino Acid Substitution within the Paramyxovirus Sendai Virus Nucleoprotein Is a Critical Determinant for Production of Interferon-Beta-Inducing Copyback-Type Defective Interfering Genomes.J Virol. 2018 Feb 12;92(5):e02094-17. doi: 10.1128/JVI.02094-17. Print 2018 Mar 1. J Virol. 2018. PMID: 29237838 Free PMC article.
-
Defective Influenza A Virus RNA Products Mediate MAVS-Dependent Upregulation of Human Leukocyte Antigen Class I Proteins.J Virol. 2020 Jun 16;94(13):e00165-20. doi: 10.1128/JVI.00165-20. Print 2020 Jun 16. J Virol. 2020. PMID: 32321802 Free PMC article.
-
CaMKII-dependent non-canonical RIG-I pathway promotes influenza virus propagation in the acute-phase of infection.mBio. 2025 Jan 8;16(1):e0008724. doi: 10.1128/mbio.00087-24. Epub 2024 Nov 27. mBio. 2025. PMID: 39601535 Free PMC article.
-
Standing on three legs: antiviral activities of RIG-I against influenza viruses.Curr Opin Immunol. 2016 Oct;42:71-75. doi: 10.1016/j.coi.2016.05.016. Epub 2016 Jun 16. Curr Opin Immunol. 2016. PMID: 27318973 Review.
Cited by
-
Preparation and Antigenic Site Identification of Monoclonal Antibodies against PB1 Protein of H9N2 Subtype AIV.Vet Sci. 2024 Sep 5;11(9):412. doi: 10.3390/vetsci11090412. Vet Sci. 2024. PMID: 39330791 Free PMC article.
-
Transient RNA structures cause aberrant influenza virus replication and innate immune activation.Sci Adv. 2022 Sep 9;8(36):eabp8655. doi: 10.1126/sciadv.abp8655. Epub 2022 Sep 9. Sci Adv. 2022. PMID: 36083899 Free PMC article.
-
Cell type- and replication stage-specific influenza virus responses in vivo.PLoS Pathog. 2020 Aug 13;16(8):e1008760. doi: 10.1371/journal.ppat.1008760. eCollection 2020 Aug. PLoS Pathog. 2020. PMID: 32790753 Free PMC article.
-
Regulation of RIG-I-like receptor-mediated signaling: interaction between host and viral factors.Cell Mol Immunol. 2021 Mar;18(3):539-555. doi: 10.1038/s41423-020-00602-7. Epub 2021 Jan 18. Cell Mol Immunol. 2021. PMID: 33462384 Free PMC article. Review.
-
Cytoplasm and Beyond: Dynamic Innate Immune Sensing of Influenza A Virus by RIG-I.J Virol. 2019 Apr 3;93(8):e02299-18. doi: 10.1128/JVI.02299-18. Print 2019 Apr 15. J Virol. 2019. PMID: 30760567 Free PMC article. Review.
References
-
- Kato H, Takeuchi O, Sato S, Yoneyama M, Yamamoto M, Matsui K, Uematsu S, Jung A, Kawai T, Ishii KJ, Yamaguchi O, Otsu K, Tsujimura T, Koh CS, Reis e Sousa C, Matsuura Y, Fujita T, Akira S. 2006. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 441:101–105. doi:10.1038/nature04734. - DOI - PubMed
-
- Shaw ML, Palese P. 2013. Orthomyxoviridae, p 1151–1185. In Knipe DM, Howley PM (ed), Fields Virology, 6th ed, vol I Lippincott Williams & Wilkins, Philadelphia, PA, USA.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
