

Review
doi: 10.1038/s41556-018-0223-z.
Epub 2019 Jan 2.
Integrin trafficking in cells and tissues
Affiliations
- PMID: 30602723
- PMCID: PMC6597357
- DOI: 10.1038/s41556-018-0223-z
Item in Clipboard
Review
Integrin trafficking in cells and tissues
Nat Cell Biol.
2019 Feb.
Abstract
Cell adhesion to the extracellular matrix is fundamental to metazoan multicellularity and is accomplished primarily through the integrin family of cell-surface receptors. Integrins are internalized and enter the endocytic-exocytic pathway before being recycled back to the plasma membrane. The trafficking of this extensive protein family is regulated in multiple context-dependent ways to modulate integrin function in the cell. Here, we discuss recent advances in understanding the mechanisms and cellular roles of integrin endocytic trafficking.
Conflict of interest statement
The authors declare that they have no financial and non-financial competing interests.
Figures
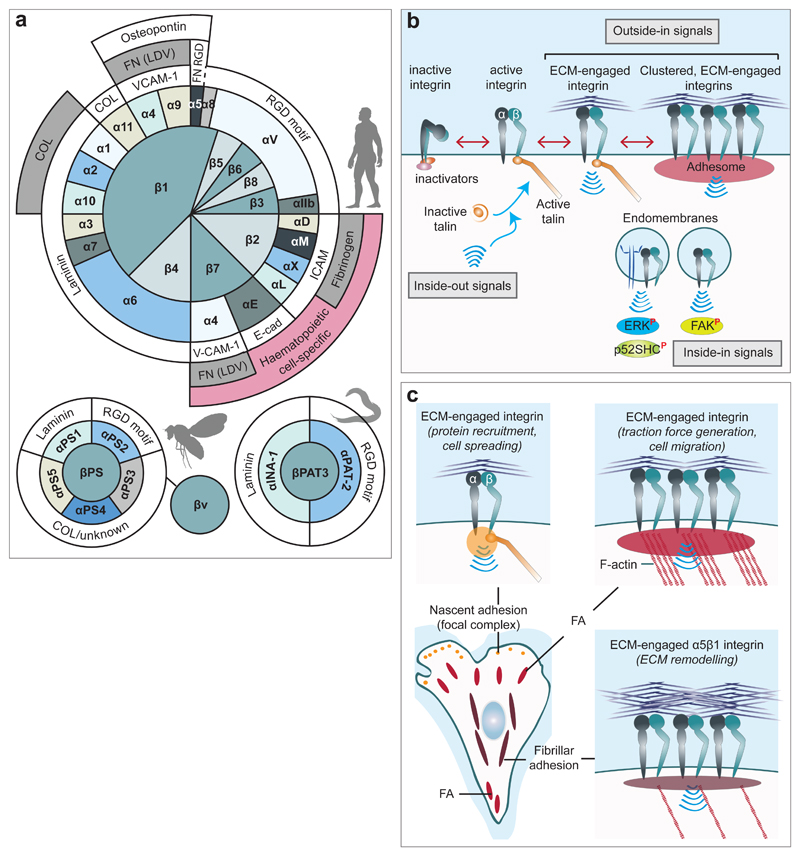
a, The reported pairing between integrin α and β
subunits and the ECM ligand/s for each heterodimer are illustrated for mammals,
Drosophila and C. elegans. In mammals,
α5β1-integrin binds to the fibronectin RGD motif, whereas
α4β1 binds to the fibronectin LDV motif. COL: collagen; E-cad:
E-cadherin; FN: fibronectin; ICAM: intercellular cell adhesion molecule; VCAM-1:
vascular cell adhesion molecule 1. For a more comprehensive/exhaustive list of
integrin ligands, please refer to. b, Integrins are bidirectional signalling molecules. Inside-out
signals regulate talin binding to integrin β-tails and thus tightly
control integrin affinity for ECM ligands. Subsequent ECM binding triggers
recruitment of protein complexes (scaffolding and adaptor proteins, kinases and
phosphatases, etc.) to the integrin cytoplasmic tails to promote integrin
downstream signalling (outside-in signalling). Integrins can also signal from
within endosomes (inside-in signalling) to support FAK activity and suppress
anoikis or to promote signalling
downstream of co-trafficking MET to support anoikis resistance, tumour growth
and cancer cell dissemination to lungs.
The superscripted P in red indicates phosphorylation. ERK, extracellular
signal-regulated kinase; p52SHC, p52 isoform of SHC-transforming protein 1. c, Integrin-ECM adhesions, in vitro, are defined
based on localisation, components and maturation stage. Nascent adhesions (focal
complexes) represent initial integrin receptor clustering in response to ECM
engagement and recruitment of adaptor and signalling proteins to the integrin
tails. These small protein assemblies mature into focal adhesions (FA), which
serve to anchor actin stress fibres and are vital for generation of contractile
force. Fibrillar adhesions are mature α5β1-integrin adhesions, and
prominent sites of fibronectin fibrillogenesis, that result from the centripetal
translocation of this specific integrin heterodimer towards the cell body.
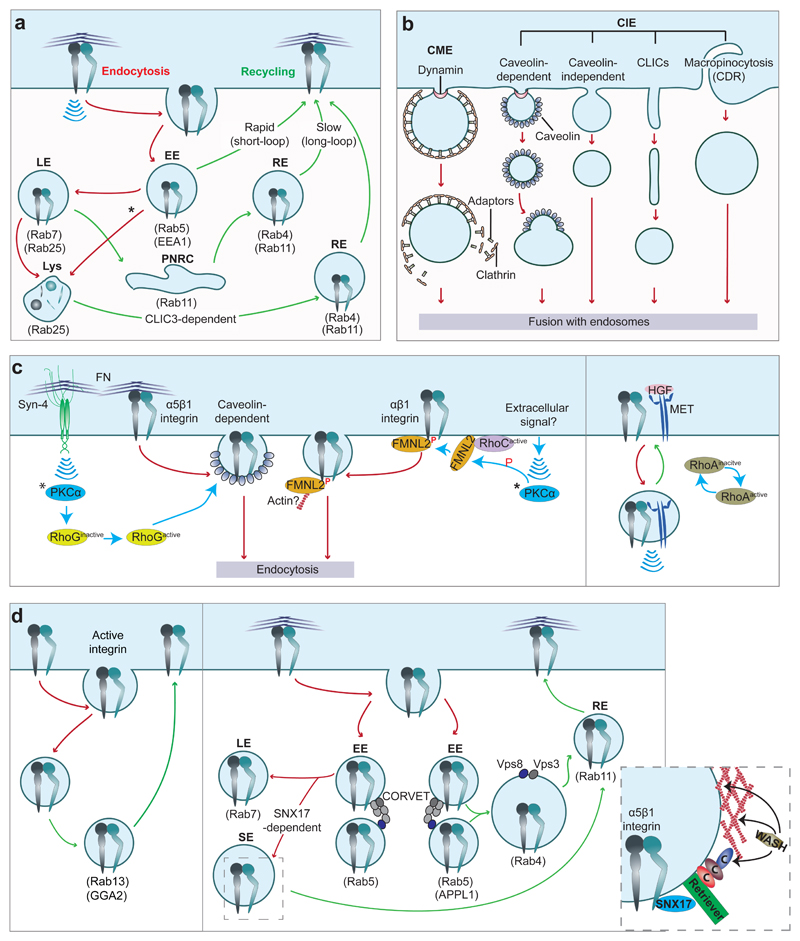
a, Summary of integrin trafficking pathways. Integrins are
internalised into Rab5-positive early endosomes (EE). EE mature into late
endosomes (LE), which fuse with lysosomes (Lys) for integrin degradation. Under
certain conditions (Rab25 and chloride intracellular channel protein 3 (CLIC3)
expression), integrins exit LE/Lys compartments and are recycled back to the
plasma membrane. Integrins also traffic
to multivesicular bodies (highlighted by *) and to the perinuclear recycling
compartment (PNRC). This process occurs within 20 minutes, while degradation
takes several hours resulting in the
majority of integrins being recycled back to the cell surface. Each step
requires a spatiotemporal hierarchy of interactions between integrins, endocytic
adaptors, Ras and Arf GTPase family members and other molecules. The Rab GTPases
involved are indicated. b, Integrin receptor internalisation mechanisms, broadly classified
as clathrin-mediated endocytosis (CME) or clathrin-independent endocytosis (CIE,
including caveolin-dependent pathways, micropinocytosis and clathrin-independent
carriers (CLICs)). CDR, circular dorsal ruffles. c, Integrin endocytosis can be fine-tuned by extracellular-initiated
signals. Left-hand panel: Syndecan-4(Syn-4)─fibronectin interaction
activates PKCα and RhoG and promotes caveolin-dependent
α5β1-integrin endocytosis, attenuated adhesion and increased cell
migration. PKCα also
phosphorylate FMNL2 (recruited by RhoC). Phospho-FMNL2 (superscripted P in red
indicates phosphorylation) interaction with the α-integrin GFFKR motif
drives αβ1-integrin internalisation. The role of actin nucleation and the extracellular
signal regulating this pathway remain undefined. *PKCα activation occurs
at the plasma membrane. Right-hand panel: MET receptor─β1-integrin
interaction induces integrin endocytosis, collective mesenchymal cell migration,
in a HIP1- (clathrin adaptor) and RhoA-dependent manner, and “inside-in” signalling (see Fig.
1b). d, Recruitment of specific endosomal adaptors imposes selectivity to
integrin recycling pathways. Left-hand panel: GGA2, an Arf effector has been
implicated in Rab13-mediated recycling of β1-integrin (a non-peer-reviewed study). Right-hand
panel: Different populations of Rab5-positive EE, defined by the presence of
Rab5 effectors EEA1 (not shown), APPL1 and the CORVET complex promote different
recycling routes,,. CORVET
mediates fission and fusion of EE that mature into LE. Here, Rab5 is replaced by
Rab7 and integrins enter LE. Integrins escape degradation by interaction with
SNX17 (components in inset)–. CORVET
components Vps3/Vps8 localise to Rab4-positive endosomes (fusion of
APPL1-positive EE). Vps3/Vps8 vesicles deliver integrins to the plasma membrane
through Rab11-positive recycling endosomes. SE, sorting endosomes.
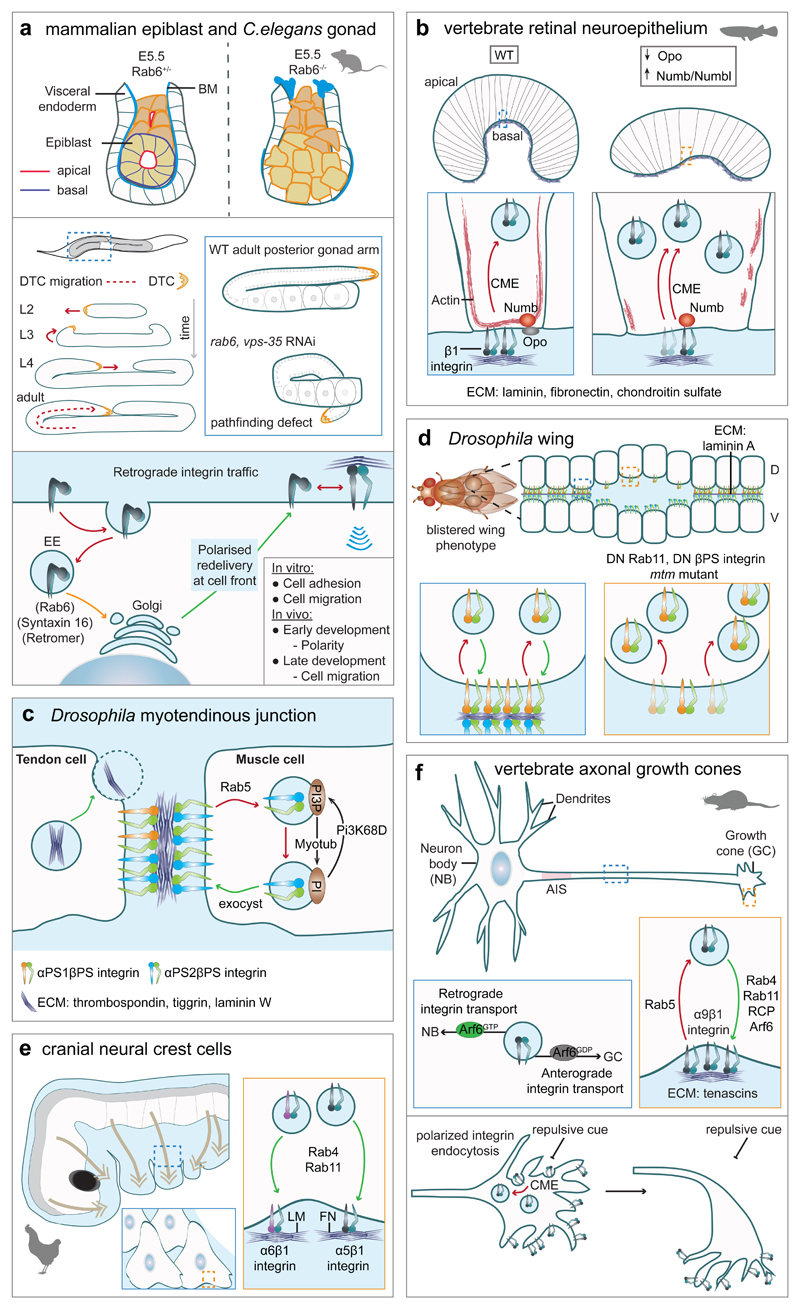
a, Retrograde transport of inactive β1-integrin (bottom) is
required for polarized cell behaviours during development. Epiblast cell rosette
structure fails to form in Rab6 mutant embryos (top). Integrin retrograde traffic is also required for
persistent migration of a distal tip cell (DTC) in C. elegans
gonad (middle). Knockdown of Rab6 or the retromer component Vps35 causes a DTC
pathfinding error and morphogenetic defects. b, Integrin CME during optic cup formation is facilitated by binding
of Numb and Numb-like to the β1-integrin NPxY motif and is inhibited by
competitive binding of these adaptors to the NPxF motif of the membrane protein
Ojoplano (Opo). Loss of Opo or overexpression of adaptors
result in excess integrin endocytosis, decreased cortical actin in basal
endfeet, failure in basal constriction and a flat retina,. c, In MTJ, the muscle cell (αPS2βPS-integrins) is
attached to the tendon cell (αPS1βPS- and
αPS2βPS-integrins) indirectly through ECM. Integrin turnover in
MTJ is increased by reduced availability of ECM and decreased by raised muscle
tension, elevated integrin outside-in activation, or expression of Rab5
mutants,. Cleavage of PI(3)P by myotubularin (MTM1) prevents
receptor accumulation in endosomal-related inclusions. d, Cellular layers forming Drosophila wing are held
together by adhesion of αPS1βPS-integrin (dorsal, D) and
αPS2βPS-integrin (ventral layer, V) to the ECM secreted in
between. In wing imaginal disc, βPS-integrin trafficking is mediated by
Rab11, which when disrupted leads
to increased apical cell area, intracellular βPS-integrin accumulation
and disorganised actin cytoskeleton. The ensuing change in cell shape from
columnar to cuboidal (not shown) leads to separation of cell layers and
blisters. DN, dominant negative; mtm mutant, myotubularin mutant. e, Recycling of α5β1- and α6β1-integrins
is required for migration of cranial NCC. Integrins are recycled through Rab4 and Rab11 pathways in a
laminin substrate-dependent manner. f, α9β1-integrin recycling within the growth cone and
long-range axonal integrin traffic are required for efficient axon growth.
Rab5-regulated endocytosis is followed by recycling through Rab4, Rab11 or Arf6
pathways. Activation of Arf6 promotes retrograde transport of
integrin-containing vesicles towards the neuron body. Chemorepellent cues trigger β1-integrin
endocytosis on one side of the growth cone, which changes direction of growth
cone migration.
References
-
- Maartens AP, Brown NH. Current Topics in Developmental Biology. Vol. 112. Academic Press; Cambridge: 2015. Anchors and Signals: The diverse roles of integrins in development; pp. 233–272. 2015. - PubMed
-
- Mould AP, Humphries MJ. Regulation of integrin function through conformational complexity: Not simply a knee-jerk reaction? Curr Opin Cell Biol. 2004;16:544–551. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
